

Evolution of the marine biological carbon pump and geological events in the Ross Sea during Modern and the Last Glacial Maximum Period—Stratigraphic records of molecular biomarkers
-
摘要: 海洋生物碳泵(BCP)与古气候的关系是碳循环研究的一个核心科学问题。 本文利用罗斯海沉积物总有机碳(TOC)和一些重要的类脂化合物所隐含的生态学特性,即那些可引起海底沉积碳库组成和海洋BCP(包括生物泵BP和微型生物碳泵MCP)效率改变的分子生物标志物,从罗斯海现代海洋出发,追溯到末次冰期以来(27~0.6 ka BP)古海洋生物碳泵演化(重点讨论BP和MCP)及其与一些重大地质事件的联系。研究结果表明:(1)罗斯海表层沉积物中相对高的TOC和较低C/N比值、以及正构烷烃色谱峰型、主峰碳、分子组合特征低碳数和高碳数比值(L/H)和(nC21 + nC22)/(nC28 + nC29)比值、低碳烃(nC15 + nC17 + nC19)、中碳烃(nC21 + nC23 + nC25)和高碳烃(nC27 + nC29 + nC31)、细菌(BrGDGT)和较低的陆源土壤指标(BIT)一致性表征海源有机质贡献占主导地位,通过海洋BP和MCP效应将内源有机碳输送到海底进行长期储存;Pr/Ph比值结合Ph/nC18和Pr/C17比值均表明表层沉积物为还原−强还原性的缺氧环境,有利于海底储存有机碳,同时还与海冰消融关系密切。(2)利用低温校正公式
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST[式(2)]反演现代的海洋表层温度(SST),并与WOA-SST(3月)表层水体温度较接近(R2 = 0.78,p < 0.01,n = 15);同时利用$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST-2重建罗斯海JB03岩芯末次冰期以来的古海洋温度在−0.74~2.86℃范围(平均为1.03℃),接近现代南极罗斯海夏季温度。(3)JB03岩芯记录年代为27.27~0.6 ka BP,分为末次冰盛期(27.3~21 ka BP)、末次冰消期(21~11.7 ka BP)和全新世(11.7~0.6 ka BP)3个地质历史时期。末次冰期古海洋受冰盖和海冰限制作用的影响,初级生产力低下、沉积速率仅为0.45 cm/ka、Pr/Ph比值、Ph/nC18和Pr/nC17反映沉积环境氧化性较强、不利于MCP和BP储碳,在寒冷气候时段碳储量潜力降低;进入全新世暖期,冰架退缩解体,温暖的气候条件有利于浮游植物生产力和硅藻增加、浮游动物增加及影响粪便物质组成和提高沉降速率,促进有机碳向深海的输送加快,海底沉积为弱还原−弱氧化环境,有利于碳保存;硅藻生物量提高就意味着硅质泵加强,进而微生物活性增强、促进古菌和细菌生长,因而微生物总量-GDGTs、 产甲烷古菌或广古菌-GDGT-0、奇古菌-Crenarchaeol生物量大大提高,显示罗斯海全新世以来古海洋BP和MCP作用大大加强,且古海洋BP与现代海洋均以硅藻/硅质泵为绝对主导。研究还发现,罗斯海末次冰期和全新世以来的古海洋BP和MCP储碳效率变化均与古海洋地质事件,即全球性大尺度的气候变化有关,这个碳库的大小与气候冷暖之间存在对应关系,罗斯海古海洋调节大气CO2的能力,尤其在全新世暖期最强,这对于认识全球气候变化的海洋调控机制具有重要的科学意义。-
关键词:
- 海洋初级生产力 /
- 生物泵 /
- 微型生物碳泵 /
- 分子生物标志物 /
- ${\mathrm{TEX}}_{86}^{{\mathrm{L}}} $
Abstract: The relationship between the marine biological carbon pump(BCP) and paleoclimate is a central scientific issue in carbon cycle research. This study utilizes the ecological characteristics implied by total organic carbon (TOC) and key lipid compounds in Ross Sea sediments, specifically molecular biomarkers that can influence the composition of the seabed carbon reservoir and the efficiency of the marine BCP (including the biological pump BP and the microbial carbon pump MCP), tracing the evolution of the ancient marine biological carbon pump (with a focus on BP and MCP) since the Last Glacial Maximum (27−0.6 ka BP) and its connection to significant geological events. The research findings indicate: (1) the relatively high TOC and low C/N ratios in surface sediments of the Ross Sea, along with characteristics such as normal alkane chromatographic peak shapes, dominant carbon, molecular composition features with low to high carbon number ratios (L/H) and(nC21 + nC22)/(nC28 + nC29)ratios, low n-alkanes(nC15 + nC17 + nC19), mid-chain alkanes(nC21 + nC23 + nC25), high-chain alkanes (nC27 + nC29 + nC31), bacterial (BrGDGT), and lower terrigenous input index (BIT), collectively indicate a predominance of marine-derived organic matter contributing to the seabed through the effects of the marine BP and MCP, facilitating long-term carbon storage. The Pr/Ph ratio combined with Ph/nC18 and Ph/nC17 ratios suggest that surface sediments represent a reducing to strongly reducing anoxic environment conducive to organic carbon sequestration on the seafloor, closely linked to sea ice melt. (2) By employing a low-temperature correction formula$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST [(Eq. (2)] to infer modern sea surface temperatures (SST) in the ocean and comparing them to WOA-SST (March) surface water temperatures, a close match is observed (R2 = 0.78, p < 0.01, n = 15). Additionally, using$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST-2 to reconstruct paleoceanic temperatures in the Ross Sea Core JB03 since the Last Glacial Maximum yields a range of −0.74℃ to 2.86℃ (average 1.03℃), approximating modern summer temperatures in the Antarctic Ross Sea. (3) The Core JB03 record spans 27.27 to 0.6 ka BP, divided into three geological periods: the Last Glacial Maximum (27.3~21 ka BP), the deglaciation period (21−11.7 ka BP), and the Holocene (11.7−0.6 ka BP). During the Last Glacial Maximum, the ancient ocean was influenced by ice cover and sea ice constraints, resulting in reduced primary productivity, sedimentation rates of only 0.45 cm/ka, and indicators such as Pr/Ph ratios, Ph/nC18, and Ph/nC17 reflecting a strongly oxidative sedimentary environment unfavorable for MCP and BP carbon sequestration, leading to diminished carbon storage potential during cold climate periods. As the Holocene warm period commenced, ice shelves retreated and disintegrated, creating favorable warm climate conditions for increased phytoplankton productivity, diatom abundance, zooplankton proliferation, alterations in fecal matter composition, and enhanced sedimentation rates, accelerating the transfer of organic carbon to the deep sea. Seabed sediments transitioned to weakly reducing to weakly oxidizing conditions, conducive to carbon preservation. Elevated diatom biomass signifies strengthened siliceous pump activity, subsequently enhancing microbial activity, promoting the growth of archaea and bacteria, leading to significant increases in total microbial-GDGTs, methane-producing archaea or widespread archaea-GDGT-0, and crenarchaeol archaeal biomass, indicating a substantial enhancement in ancient marine BP and MCP activity in the Ross Sea since the Holocene, with diatoms/siliceous pumps dominating both ancient and modern marine environments. The study further reveals that changes in carbon sequestration efficiency of the ancient marine BP and MCP in the Ross Sea during the Last Glacial Maximum and the Holocene are linked to geological events, specifically large-scale global climate variations, where the size of this carbon reservoir corresponds to climatic fluctuations. The Ross Sea’s ability to regulate atmospheric CO2 through ancient marine processes, particularly during the Holocene warm period, holds significant scientific implications for understanding the oceanic regulatory mechanisms of global climate change. -
1. 引言
地球上最活跃的碳库是海洋。 海洋生物碳泵(Biological Carbon Pump, BCP),包括生物泵(Biological Pump, BP)和微型生物碳泵(Microbial Carbon Pump, MCP),是海洋生态系统通过碳循环调节地球环境变化的关键途径之一[1]。在一个相对稳定的时期里,BP和MCP对气候变化的影响举足轻重[2]。经典生物泵(BP)是一个至关重要的沉降(竖向)储碳机制[3],BP主要启动于真光层的光合作用,通过浮游植物自身以及在食物链传递过程中将 CO2 固定为颗粒有机碳(Particulate Organic Carbon, POC),再通过一系列的BP 机制,使得一部分POC可以到达海底沉积物、不再进入碳循环、从而实现长期储碳[4]。MCP是一个基于溶解有机碳的非沉降(横向)储碳机制[5–6],时刻影响着海洋碳循环,MCP通过微型生物的作用将活性的溶解有机碳(Dissolved Organic Carbon, DOC)转化为生物难以利用的惰性有机碳(Recalcitrant Dissolved Organic Carbon, RDOC)使得该部分碳进入水体长期积累,构成海洋水柱储碳,这就是微型生物碳泵[7]。海洋水柱的碳封存,部分通过黏土矿物等沉降到沉积物里,构成了海洋内部碳的长期储存[8]。全球气候变化,加强了海洋MCP的研究[9–10]。当下,我国科学家提出的MCP理论,开辟了海洋碳汇研究的新领域,服务于碳中和战略目标。
浮游动植物是海洋主要生产者,也是现代生物泵的重要组成部分,初级生产力的微小变化,也会影响二氧化碳的浓度。目前国内外利用沉积物类脂生物标志物反演浮游植物生产力、群落结构和浮游动物总量,已得到了较好的应用[11–15]。微生物细胞膜脂的甘油二烷基甘油四醚化合物(Glycerol Dialkyl Glycerol Tetraethers, GDGTs)属于大分子生物标志化合物,特定的GDGTs 可能是海洋沉积物RDOC库的重要组成部分[16],GDGTs 作为特定的地质微生物标志物,在各种环境中广泛存在,如厌氧的产甲烷古菌(GDGTs-0)和好氧的氨氧化奇古菌(GDGTs-4)的分布特征能够敏感地响应环境的变化[17–19]。特定的GDGTs 分子指标在重建海水温度、营养盐、pH、氧化还原状况[20–25]环境参数中取得了大量研究成果,并可追溯到白垩纪和侏罗纪[26–29],但GDGTs相关研究在罗斯海仍缺乏。
南大洋是典型碳汇动态特征的海域,对全球碳汇贡献作用显著[30],且与大洋生物碳泵(BP和MCP)存在着紧密联系[31–33],罗斯海是太平洋南端的一个最高纬度边缘海,具有典型的极地环境特征,海冰区拥有特殊生态,包括鲸鱼、企鹅、海豹和南极磷虾等海洋浮游动物,生长着大量的冰藻和细菌等生物[34–35]。大气CO2浓度的升高,罗斯海正在发生快速的变化[36–37],国内外科学家开展了大量罗斯海生态系统和生物泵的研究[38–44],旨在了解全球变化下生物泵在南大洋生态系统中的作用和地位,同时对南极生物泵的研究也需要更长时间序列和更系统的观测来验证与校正[45]。本文对罗斯海表层沉积物和位于Joides海槽南端岩芯JB03(进行高精度14C年代序列)同步开展现代和古海洋生态及其MCP与BP研究,重建一些关键时期的全球古气候变化事件与古海洋沉积碳库变动的关联,重点分析这些时期的浮游动植物生产力/群落结构、微生物生产力、古菌类群的时空分布特征和储碳效率与环境变化(如氧化还原、海水升温)之间的联系,探究地质历史时期的海洋生产者和生物碳泵的演化,及其生态系统储碳格局与一些重大地质事件的关系,以古示今是地球科学领域思考和解决问题的重要方法,了解古海洋生产者和生物碳泵储碳(碳汇)历史演化认识,对理解现今海洋碳循环的过程和机制有重要启示意义。总体来讲,我们对于地质时期海洋生物碳泵了解很少。
2. 海域概况和样品采集
罗斯海(Ross Sea)是南大洋深入南极洲形成的一个边缘海,边缘海因其生产力高、对有机碳的埋藏贡献大等特点,对全球碳循环和碳埋藏具有重要意义。罗斯海拥有丰富的生物多样性和南极最大的浮动冰原之一,对全球气候变化非常敏感,当下这部分区域正在快速响应全球的气候变化[46]。边缘海对大气CO2的源汇作用是碳循环研究中最基础也是最重要的部分,它的强弱反映边缘海对气候变化的调节能力。
本研究所用样品是中国第 31 和 32 航次南极科学考察期间在罗斯海采集的 18 个表层沉积物(JB01、JB03、JB04、JB05、R02、R04、R05、R08、R09、R19、RB02B、RB03B、RB05B、RB06B、RB07B、RB08B、RB11B、RB16B)和 1 根 JB03 沉积柱样。表层沉积物采样站位主要位于维多利亚地陆架边缘、罗斯冰架前缘(世界上最大的冰架)、Joides 海槽和 Glomar Challenger 海槽(图1),由箱式采样器采集。表层沉积物由砂、粉砂和黏土组成,且以粉砂为主,具有显著的区域特征。JB03 岩芯(75.8°S, 170.7°E)位于罗斯海 Joides 海槽,采用重力采集。岩芯全长 131 cm,厘米级分割。所有样品现场冷冻保存后带回。
 图 1 罗斯海表层沉积物和岩芯采样站位箭头代表流向,灰色为表层环流,蓝色为变性绕极深层水(MCDW)涌入陆架Figure 1. Ross Sea surface sediment and core sampling stationsArrows represent flow direction, grey represent the surface gyre, and blue represent the Modified Circumpolar Deep Water (MCDW) spilling onto the shelf
图 1 罗斯海表层沉积物和岩芯采样站位箭头代表流向,灰色为表层环流,蓝色为变性绕极深层水(MCDW)涌入陆架Figure 1. Ross Sea surface sediment and core sampling stationsArrows represent flow direction, grey represent the surface gyre, and blue represent the Modified Circumpolar Deep Water (MCDW) spilling onto the shelf3. 实验方法
3.1 AMS14C分析方法
加速质谱仪(AMS)方法是计测样品中现存的14C原子数来确定年龄,是基于14C同位素的原子数目因放射性衰变而随时间指数衰减的规律,准确地说是测定样品中现存的14C与12C原子数比值。柱样沉积物的11个子样(层位0~2 cm、2~4、18~20、54~56、68~70、72~74、78~80、102~104、110~112、112~114、128~130 cm)使用总有机碳(TOC)的14C测定(在美国Beta碳-14生物基实验室)。加速器质谱14C测定,加速器系统使用能量250 keV进行分析NEC单级粒子,结果通过ISO/IEC-
17025 :2017认证。简单来讲,样品前处理,经过超声波混合和筛选去除杂质无机碳,保留有机碳部分,精确制备加速器质谱仪14C石墨靶系统,标准为NIST SRM-4990C草酸,进行加速器质谱仪测试。Δ14C的分析精度测量值通常小于5‰。3.2 总有机碳 TOC 和总有机氮 TN 测定
使用元素分析仪(德国Elementar Vario MICRO cube)同时测定TC和TN。TOC的测定需要先用酸去除无机碳,采用的是差减法。简而言之,将0.5 g冷冻研磨后的沉积物样品置于15 mL玻璃试管,加入10 mL (1 mol/L) HCl溶液充分搅拌混合,置于恒温50℃水浴锅中反应48 h后,样品离心分离(
2500 r/min,5 min)将上层清液倒入新容器,用蒸馏水清洗直至pH值达到中性,最后样品冷冻干燥并称重;准确称重脱碳样品[(30.0 ± 0.2) mg]置于锡胶囊中进行分析,标准偏差小于1%。3.3 生物标志物提取与分离
准确称重 5 g 冻干后研磨的沉积物样品,装入快速溶剂萃取仪(ASE-350, 美国)中,加入氘代正二十四烷(nC24D50)、C46-GDGTs 和正十九醇(19-OH)作为内标;用二氯甲烷和甲醇(V∶V,9∶1)在温度 100℃加热 5 min 条件下,静态萃取 10 min,循环提取 3 次,萃取液经旋转蒸发后氮吹仪浓缩处理,转移到 10 mL 玻璃瓶后吹干;加入 6%氢氧化钾−甲醇溶液超声 10 min,重复超声1次,置于干燥器中避光水解 12 h;取出水解液超声 10 min,加入 4 mL 正己烷后涡旋振荡,放置分层后将上层清液转移到 20 mL 玻璃瓶,重复萃取 4 次,合并所需的有机质组分(氮吹仪吹至 0.2 mL);用湿法硅胶柱分离总脂类,经正己烷淋洗获得非极性组分,再经二氯甲烷:甲醇(V∶V,1∶1)淋洗得到极性组分。其中非极性组分用于分析测定正构烷烃,极性组分中的一部分用氮吹仪吹干,加入少量正己烷:异丙醇(V∶V,95∶5)溶液,过0.45 μm 聚四氟乙烯滤膜(PTFE)滤膜,用于分析测定 GDGTs;另一部分用双(三甲基硅烷基)三氟乙酰胺(BSTFA)衍生化试剂,在 70℃恒温下反应 1 h,衍生化后分析测定甾醇和长链烯酮。
3.4 生物标志物测定与分析
正构烷烃:用安捷伦气相色谱(Agilent 6890N,美国)进行测试分析,色谱柱为 HP-1Methy1Siloxane(50 m × 0.32 mm × 0.17 μm)。分析测试条件:进样温度:310℃,FID 检测器温度:320℃。采用不分流进样,载气(N2)流速:1.2 mL/min。升温程序:初始温度为60℃,保持 1 min,以 10℃/min 的升温速率升至200℃,再以 5℃/min 的升温速率升至 300℃,保持 5 min,然后以 5℃/min 的升温速率升至 310℃,保持 15 min。样品的定性与定量:在相同色谱条件下,目标分子化合物的保留时间根据 34 种正构烷烃(nC7~nC40)混标的保留时间对照来判定;采用 nC7~nC40 正构烷烃混标各组分峰和 nC24D50 内标峰的相对响应值定量分析,通过目标化合物峰面积与正构烷烃混标各组分内标峰面积,计算出样品各组分含量。
甾 醇 及 长 链 烯 酮 : 用 安 捷 伦 气 相 色 谱 (Agilent 6890N,美国 ) 进 行 分 析 测 定 , 色 谱 柱 用 HP-1Methy1Siloxane(50 m × 0.32 mm × 0.17 μm)。分析测试条件:进样温度:310℃,FID 检测器温度:320℃。采用不分流进样,载气(N2)流速:1.2 mL/min。升温程序:初始温度为 80℃,保持 1 min,以 25℃/min 的升温速率升至 200℃,再以 3℃/min 的升温速率升至 250℃,然后以 1.8℃/min 的升温速率升至 300℃,保持 8 min,5℃/min 升至 310℃,保持 5 min。甾醇的气相谱图(如图2b 所示)。极性组分的目标分子化合物根据相同条件下 GC 和 LC-MS 特征离子峰和保留时间并结合相关文献来确定目标化合物;采用内标法定量分析,通过目标峰与内标(19-OH、nC46-GDGT)峰面积的比值来确定各目标化合物的含量。
 图 2 表层沉积物(a)正构烷烃(RB08B、JB03和RB06B)和(b)甾醇(JB01)气相及(c)GDGTs(JB01)液相色谱图Figure 2. Gas (a) and liquid (c) chromatograms of the surface sediments for (a) n-alkanes (RB08B, JB03, and RB06B) and (b) sterols (JB01) and (c) GDGTs (JB01)
图 2 表层沉积物(a)正构烷烃(RB08B、JB03和RB06B)和(b)甾醇(JB01)气相及(c)GDGTs(JB01)液相色谱图Figure 2. Gas (a) and liquid (c) chromatograms of the surface sediments for (a) n-alkanes (RB08B, JB03, and RB06B) and (b) sterols (JB01) and (c) GDGTs (JB01)甘油双烷基甘油四醚(GDGTs):利用超高效液相色谱质谱联用仪(Waters Acquity UPLC Xevo TQMS,美国)分析测定,液相色谱柱(BEH HILIC 2.1×5 mm与 2.1×150 mm,1.7 μm 串联),柱温保持 30℃,进样体积 5 μL。分析测试条件[28]:用正己烷(A)和正己烷:异丙醇(V∶V,1∶9)(B)作为流动相,洗脱梯度为 0~20 min,82.0%A:18.0%B;20.0~40.0 min,A 由 82.0%线性递减至 65.0%,B 由 18.0%线性递增至 35.0%;40.0~60.0 min 期间,A 由 65.0%线性递减至 0.0%,B 由 35.0%线性增加至 100.0%,保持 5 min;再迅速冲洗色谱柱 A 恢复到 82.0%,B 恢复到 18.0%,流速为 0.2 mL/min,保持 20 min。化合物在大气压化学电离源(APCI)中进行离子化,电晕电压为 1.5 kV,电晕电流 2.20 μA,锥孔电压 33 V,提取器电压 3 V,离子源温度为 150℃,探针温度为 550℃,锥孔气体流速 100 L/h,脱溶剂气体流速
1000 L/h,碰撞气体流速 0.15 mL/min。本次实验采用单离子检测(Single Ion Monitoring,SIM)的模式进行扫描,扫描离子质核比(m/z)为1302 、1300 、1298 、1296 、1292 、1050 、1048 、1046 、1036 、1034 、1032 、1022 、1020 、1018 ,得到 GDGTs 的液相色谱−质谱图甘油双烷基甘油四醚(GDGTs):利用超高效液相色谱质谱联用仪(Waters Acquity UPLC Xevo TQMS,美国)分析测定,液相色谱柱(BEH HILIC 2.1×5 mm 与 2.1 × 150 mm,1.7 μm 串联),柱温保持 30℃,进样体积 5 μL。分析测试条件[28]:用正己烷(A)和正己烷∶异丙醇(V∶V,1∶9)(B)作为流动相,洗脱梯度为 0~20 min,82.0%A:18.0%B;20.0~40.0 min,A 由 82.0%线性递减至 65.0%,B 由 18.0%线性递增至 35.0%;40.0~60.0 min 期间,A 由 65.0%线性递减至 0.0%,B 由 35.0%线性增加至 100.0%,保持 5 min;再迅速冲洗色谱柱 A 恢复到 82.0%,B 恢复到 18.0%,流速为 0.2 mL/min,保持 20 min。化合物在大气压化学电离源(APCI)中进行离子化,电晕电压为 1.5 kV,电晕电流 2.20 μA,锥孔电压 33 V,提取器电压 3 V,离子源温度为 150℃,探针温度为 550℃,锥孔气体流速 100 L/h,脱溶剂气体流速1000 L/h,碰撞气体流速 0.15 mL/min。本次实验采用单离子检测(Single Ion Monitoring,SIM)的模式进行扫描,扫描离子质核比(m/z)为1302 、1300 、1298 、1296 、1292 、1050 、1048 、1046 、1036 、1034 、1032 、1022 、1020 、1018 ,得到 GDGTs 的液相色谱−质谱图(图2c)。4. 结果与讨论
4.1 重建现代海洋浮游生物生产力/群落结构对生物碳泵储碳(碳汇)作用
4.1.1 TOC和正构烷烃与初级生产力/生物碳泵物质来源及其环境影响
碳汇分为陆地碳汇和海洋碳汇。海洋碳汇是指通过海洋活动和海洋生物吸收大气中的CO2,并将其固定和储存在海洋中的过程和机制。初级生产作为生物泵的起点,决定了浮游植物光合作用固定 CO2 的强弱[47]。前人测定表层沉积物年代为晚全新世以来,表层沉积物参数可与现代环境对比,本次研究的罗斯海表层沉积物中TOC含量在0.40%~1.34%(平均值为0.79%)(图3a),TOC是反映海洋生产力最常见的指标[48],与周郑鹏等[49]研究的罗斯海陆架区 TOC 平均值(0.81%)和宋乐慧等研究的罗斯海西部TOC 值(0.40%~1.56%,平均值为 0.73%)相近[50],但明显低于南极半岛表层沉积物(平均为 1.05%)[51]。表层沉积物中TOC与TN含量(图3a和b)具有显著的正相关关系(R = 0.99,n = 18,p <0.01)(图4a),表明TOC与TN具有相同来源,也意味着海洋储碳与氮循环具有密切的耦合关系。沉积物中TOC/TN比值变化范围为4.90~5.99(平均5.53)(图3c),海源有机质TOC/TN比值5~8,陆源大于12[52],指示了以海源生物碳汇为主,TOC与本区域黏土及平均粒径均无相关性(图4b),说明表层沉积物中的黏土含量不是控制有机碳分布的主要因素。罗斯海表层沉积物的TOC与TN两者含量随着地理变化呈现出显著距离减少模式,西高东低的分布趋势,这种分布特征主要取决于初级生产力和有机质的保存条件,由于生物死亡埋藏之后所经历的后生作用和生物化学降解作用程度不同所致。当颗粒有机质(POM由生物或非生物两部分组成)在水体中沉降速率相近时,相对浅的陆架水深,边缘海往往具有更高的生物泵效率和有机质累积率[53],同时还与海冰消融关系密切[54],海冰具有显著的季节性和年际变化。每年海冰消融时,区域海面浮冰覆盖率差异大,海冰密集度呈现西部低东部高的分布特征,海冰的边缘动力学强烈影响整体浮游植物的生产量[54–55]。随着海冰融化,表层浮游植物大量旺发,罗斯海西部具有较高的海洋生物泵效率,西南部冰间湖沉积速率较高,沉积物中生物碳也相应富集,海水叶绿素 a 和初级生产均显著高于其他罗斯海区域[56];而中部和东部海域由于海冰消融较晚,中部和东部海域沉积速率效率低[57],浮游植物缺少光照,初级生产低[58]。沉降的速度慢,意味着有机物还没有沉降到海底就转化为二氧化碳了,因此不利于沉积物有机质的保存。
 图 3 a. 初级生产力-TOC含量(%),b. 总有机氮-TN含量(%),c. T/N比值Figure 3. a. Primary productivity-TOC content (%), b. total organic nitrogen-TN content (%), c. T/N ratio
图 3 a. 初级生产力-TOC含量(%),b. 总有机氮-TN含量(%),c. T/N比值Figure 3. a. Primary productivity-TOC content (%), b. total organic nitrogen-TN content (%), c. T/N ratio 图 4 a. TOC含量(%)与TN含量(%),b. TOC含量(%)与黏土含量相关性Figure 4. a. TOC content (%) vs TN content (%), b. TOC content (%) vs clay content correlation
图 4 a. TOC含量(%)与TN含量(%),b. TOC含量(%)与黏土含量相关性Figure 4. a. TOC content (%) vs TN content (%), b. TOC content (%) vs clay content correlation不同来源的海洋生物类型和陆地高等植物的碳汇意义和功效也不尽相同,一般细菌和藻类等微生物正构烷烃碳数分布范围为 nC15~nC20,主峰碳为 nC15、nC17 或 nC17,低碳烃(nC15 + nC17 + nC17)来源菌藻类[59–60];水生维管束植物(大型海藻、海草等)正构烷烃碳数分布范围为,以 nC21、nC23 或 nC25 为主峰,中碳烃(nC21 + nC23 + nC25)来源大型浮游植物[61];而陆生维管束植物正构烷烃碳数分布范围为,以 nC27、nC29 或 nC31具有显著的奇碳优势[62],其碳优势指数 CPI 值大于 5,高碳烃(nC27 + nC29 + nC31)来源陆地高等植物[63]。罗斯海现代表层沉积物正构烷烃总量(∑n-alkanes)493.02~
2221.40 ng/g之间,呈现出中部高,南北低的分布趋势,在Joides海槽和Glomar Challenger海槽均出现一个高值区(图5a),从正构烷烃分子组合特征来看,碳数变化范围在nC12~nC35之间,大部分站位为前峰型且主峰碳为nC17和nC19(图2a),利用气相色谱峰型和主峰碳可直接判断出海洋来源有机质大于陆源。饱和烃低碳数(ΣnC13~21)与高碳数(ΣnC22~34)的比值(L/H)作为有机质输入的指标,其比值越大就表明海源有机质输入比例越高,反之则陆源的贡献越多[64–65]。罗斯海表层沉积物中饱和烃低碳数与高碳数的比值(L/H)范围为0.38~6.72,平均值为2.32,略大于南极半岛(范围为0.5~3.14,平均为1.83)[51]。Philip和Gilbert[66]提出陆源有机质中饱和烃(nC21 + nC22)/(nC28 + nC29)的比值为0.6~1.2,而海源为1.5~5.0,罗斯海表层沉积物(nC21 + nC22)/(nC28 + nC29)的比值在0.70~6.31之间,平均值为2.18,表明海源大于陆源,且饱和烃L/H和(nC21 + nC22)/(nC28 + nC29)比值具有显著的正相关关系,R = 0.97 (n = 18,p < 0.01),均呈现西高东低的分布趋势,与TOC和TN分布趋势一致,从以上研究可推测,碳汇所处的位置及其机制。海洋自身源是罗斯海最主要的海洋生物碳库(碳汇),水体生物通过光合作用、生物地球化学过程等自然机制产生有机质,并长期储存在沉积物中。 图 5 现代表层沉积物中TOC和正构烷烃总量及其分子组合特征与海洋生物泵联系a. 正构烷烃总量(∑n-ALK),b. 菌藻类(nC15+nC17+nC19),c. 大型浮游植物(nC21+nC23+nC25),d. 陆源高等植物(nC27+nC29+nC31),e. 姥鲛烷/植烷(Pr/Ph),f. Ph/nC17值,g. Pr/nC18值,h. L/H值,i. (nC21+nC22)/(nC28+nC29)值,j. CPI值 ,k. OEP值Figure 5. Comparison of TOC and total normal alkane and its molecular composition characteristics in modern sediments with marine biological pumpa. Normal alkane (∑n-ALK) , b. fungi algae (nC15+ nC17 +nC19), c. large phytoplankton (nC21+nC23+nC25), d. terrestrial higher plants (nC27+nC29+nC31), e. cetane/phytane (Pr/Ph), f. Ph/nC17, g. Pr/nC18, h. L/H, i. (nC21+nC22)/(nC28+nC29), j. CPI, k. OEP
图 5 现代表层沉积物中TOC和正构烷烃总量及其分子组合特征与海洋生物泵联系a. 正构烷烃总量(∑n-ALK),b. 菌藻类(nC15+nC17+nC19),c. 大型浮游植物(nC21+nC23+nC25),d. 陆源高等植物(nC27+nC29+nC31),e. 姥鲛烷/植烷(Pr/Ph),f. Ph/nC17值,g. Pr/nC18值,h. L/H值,i. (nC21+nC22)/(nC28+nC29)值,j. CPI值 ,k. OEP值Figure 5. Comparison of TOC and total normal alkane and its molecular composition characteristics in modern sediments with marine biological pumpa. Normal alkane (∑n-ALK) , b. fungi algae (nC15+ nC17 +nC19), c. large phytoplankton (nC21+nC23+nC25), d. terrestrial higher plants (nC27+nC29+nC31), e. cetane/phytane (Pr/Ph), f. Ph/nC17, g. Pr/nC18, h. L/H, i. (nC21+nC22)/(nC28+nC29), j. CPI, k. OEP值得关注的是,JB03和RB06B站的 L/H(ΣnC13~21/ΣnC22~34)和(nC21 + nC22)/(nC28 + nC29)比值异常低(分别为 0.38 和 0.70),色谱图也显示出弱双峰型特征(见图2b和c),明显区别于其他站样品,具有相对高含量的高碳数饱和烃和主峰碳为 nC25和nC19的特征,这与浮游硅藻为生源的高碳数饱和烃分布特征相似,如Nichols等[67]对南极海冰硅藻研究,也表明能够产生高碳数的正构烷烃,还有许多科学家通过硅藻培养实验均表明硅藻可以产生无奇偶优势的长链饱和烃[11, 68]。JB03 站附近所处表层环流(沿岸流)向西流时,受到 Mawson 浅滩的阻挡,经 Joides 海槽向外流分支与涌上陆架的南极 MCDW混合流速较缓[69],使得该区域水体稳定性好,有利于浮游植物的生长[58]和生物有机碳的沉积保存、意味着生物碳汇效率较高(本次测定TOC 含量为 0.91%),这与 DeMaster 等提出的14C 测定沉积速率较高的沉积物中有机质的保存效率相对较高的说法[70]相一致。罗斯海表层沉积物反演菌藻类的低烃分子含量(nC15 + nC17 + nC19)为 63.78~655.36 ng/g,来源于大型浮游植物的中碳烃含量(nC21 + nC23 + nC25)为 75.30~409.59 ng/g,低烃分子含量略大于大型浮游植物的中碳烃含量(图5b和c)。科学调查发现,不同群落的浮游植物对环境变化具有显著的反馈作用,一旦硅藻等较大粒径浮游植物占统治地位时,生态系统以 BP 为主导实现碳的捕获[71]。
沉积物的碳赋存还主要取决于氧化还原环境[10]。沉积物有机质除了最具反应性的部分之外,氧化性好,更多的有机质降解得到再循环,而在缺氧情况下有机质降解缓慢,沉积物储碳效率加强[72]。地质体中的姥鲛烷(Pristan,Pr)和植烷(phytane,Ph)主要来源叶绿素,叶绿素中的植醇广泛存在于海洋环境中,在氧化-弱氧化条件下容易发生氧化反应,进而脱羧转化成姥鲛烷,而在还原条件下则加氢发生脱水反应转化成植烷[73]。姥鲛烷/植烷(Pr/Ph)是指示沉积环境氧化还原性的重要指标,当Pr/Ph < 1时,指示还原−强还原的缺氧沉积环境;Pr/Ph > 1时,指示弱氧化−氧化环境[74],在1.0~3.0之间则指示一种贫氧的沉积环境[75]。罗斯海表层沉积物中Pr/Ph为0.39~0.93(平均为0.59),呈西南低东北高的分布特征(图5e),与海冰分布特征基本一致[56]。姥鲛烷和植烷分别与相邻饱和烃nC17、nC18的比值(Pr/nC17,Ph/nC18)亦可指示海洋生态沉积环境,当Pr/nC17 > 1时,沉积有机质中浮游藻类输入较多,罗斯海表层沉积物中Pr/nC17比值均小于0.53(图5f);Ph/ nC18 > 1时,则古菌的贡献较多,且比值越大表明沉积环境还原性越强[76],表层沉积物中Ph/ nC18的比值在0.78~1.90之间1(图5g),平均值为1.1,进一步佐证研究海区为还原−强还原性的缺氧沉积环境,生物降解作用较弱,有利于海洋生物碳泵(BP和MCP)海底储存有机碳。
饱和烃的碳优势指数(Carbon Preference Index,CPI)的值可用于指示有机质早期成岩作用[77],早期成岩作用是埋藏在海底的沉积物以有机质的降解为驱动力,与周围水环境相互作用而发生的一系列物理、生物、化学变化[78]。罗斯海表层沉积物CPI = 1/2(C25 + C27 + C29 + C31 + C33 + C35)/(C24 + C26 + C28 + C30 + C32 + C34)+ (C25 + C27 + C29 + C31 + C33 + C35)/(C26 + C28 + C30 + C32 + C34 + C36)和OEP = (C27 + 6C29 + C31)/(4C28 + 4C30)呈现低值,变化范围为1.01~2.43(平均1.66)和 1.29~3.27(平均 2.02),无明显的奇偶优势(图5j),表明研究海区沉积物中有机质演化程度较低,呈现出相对高含量的无奇偶优势高碳数正构烷烃,可以推测这是Terra Nova Bay和罗斯海南端的冰间湖受海域硅藻旺发[58, 79]所贡献的高碳烃正构烷烃的结果[11, 67, 80]。
4.1.2 GDGTs与微生物总量/古菌类群及其海洋MCP储碳作用
MCP储碳是海洋生态系统中一系列微型生物活动的总体效应[5],核心驱动力是一类特殊功能的微型生物类群(细菌/古菌等)将活性溶解有机碳(LDOC)高效地转化为惰性的溶解有机碳(RDOC),进而起到储碳的过程,RDOC库的积累可以缓解气候变化。GDGTs是微生物细胞膜脂的重要组成部分,是惰性 RDOC 的主要贡献者。MCP则通过微型生物的作用将活性溶解有机碳(DOC)转化为惰性有机碳(RDOC),从而增加了碳在海洋中的停留时间[5]。罗斯海大部分站表层沉积物中GDGTs含量为87.93~828.83 ng/g(除JB01站显著高于其他站外),整体与普里兹湾和南极半岛附近海域结果相近,呈现西高东低的分布趋势(图6a),与TOC、正构烷烃分子组合、菌藻类(nC15 + nC17 + nC19)、大型浮游植物(nC21 + nC23 + nC25)、L/H和(nC21 + nC22)/(nC28 + nC29)比值(海洋来源占优势)分布相一致(图3a和图5a、b、c、h和i)。GDGTs从结构上可分为类异戊二烯(Isopernoidal GDGTs,简称isoGDGTs)和支链(Branched GDGTs,简称brGDGTs)两大类,细胞膜脂 brGDGTs 和 isoGDGTs 作为特定分子生物标志物被认为是土壤细菌和海洋古菌细胞膜所产生,可反应细菌和古菌生物量[81–82]。
 图 6 现代表层沉积物微生物细胞膜脂GDGTs、不同类群的古菌及其与MCP储碳联系a. 微生物细胞膜脂-GDGTs含量,b. 古菌-IsoGDGTs含量,c. 细菌-BrGDGTs含量,d. 产甲烷古菌-GDGT-0含量,e. 奇古菌-Crenarchaeol含量,f. R0/5值,g. BIT值Figure 6. Modern surface sedimentary microbial cell membrane GDGTs, archaeal groups and their link to MCP carbon storagea. Microbial cell membrane lipids-GDGTs content, b. Archaea-IsoGDGTs content, c. Bacteria-BrGDGTs content, d. Methanogenic archaea-GDGT-0 content, e. Thaumarchaeota-Crenarchaeol content, f. R0/5 values, g. BIT values
图 6 现代表层沉积物微生物细胞膜脂GDGTs、不同类群的古菌及其与MCP储碳联系a. 微生物细胞膜脂-GDGTs含量,b. 古菌-IsoGDGTs含量,c. 细菌-BrGDGTs含量,d. 产甲烷古菌-GDGT-0含量,e. 奇古菌-Crenarchaeol含量,f. R0/5值,g. BIT值Figure 6. Modern surface sedimentary microbial cell membrane GDGTs, archaeal groups and their link to MCP carbon storagea. Microbial cell membrane lipids-GDGTs content, b. Archaea-IsoGDGTs content, c. Bacteria-BrGDGTs content, d. Methanogenic archaea-GDGT-0 content, e. Thaumarchaeota-Crenarchaeol content, f. R0/5 values, g. BIT values罗斯海表层沉积物中的brGDGT平均含量为17.99 ng/g,其中GDGT-Ⅲa平均含量为5.94 ng/g和GDGT-Ⅱb平均含量为6.94 ng/g,略高于其他类型(GDGT-Ⅲc、GDGT-Ⅱb、GDGT-Ⅰc),表层沉积物中细菌−brGDGTs含量很低(图6c),占GDGTs总量的3%左右,brGDGTs与TOC含量呈现良好的线性关系,这佐证了罗斯海细菌生长受有机质限制的结论,因为微生物菌要成活,碳源是细菌最重要的营养物质,是构成细菌细胞结构和代谢产物的基本原料[83]。表层沉积物中古菌-isoGDGTs为87.51~810.86 ng/g(平均419.58 ng/g)(图6b),isoGDGTs与TOC含量(图3a)具有一致分布特征,整体均呈现西高东低,其含量与冰间湖叶绿素 a 分呈现良好的线性关系[56],可能是初级生产为产生isoGDGTs古菌提供能量来源。isoGDGT的5种古菌为GDGT-0、GDGT-1、GDGT-2、GDGT-3、GDGT-4(Crenarcheaol),其中GDGT-1、GDGT-2、GDGT-3含量较低,GDGT-0(图6d)和GDGT-4(Crenarcheaol)(图6e)二者之和占isoGDGTs总量的96%以上。罗斯海表层沉积物中奇古菌醇(Crenarchaeol)含量为(37.76~953.42) ng/g小于产甲烷古菌-(GDGT-0)含量为(46.97~
1545.97 ng/g),这种状况有别于温暖地区沉积物中isoGDGTs微生物古菌类型[16],即温暖地区Crenarchaeol含量大于产甲烷古菌-GDGT-0含量。目前认为奇古菌醇-Crenarchaeol及其异构体的生物源相对简单,主要来自奇古菌;GDGT-0可能会受到多源输入的影响,不仅有产甲烷古菌的贡献[84–85],可能来源于不同类型的奇古菌、泉古菌以及广古菌[86]。基于此,有学者提出R0/5值(GDGT-0/crenarchaeol)指标评估产甲烷古菌的贡献程度,该指标的生物学基础是产甲烷古菌合成GDGT-0而无法合成crenarchaeol(而crenarchaeol可产生GDGT-0),产甲烷古菌的R0/5 > 2[87–88],海洋奇古菌的R0/5值为0.2~2.0 [20, 89],以指示奇古菌和产甲烷古菌对isoGDGTs的贡献。研究区各站沉积物中R0/5值为1.22~1.86(平均1.51)(图6f),R0/5值普遍小于2,说明罗斯海表层沉积物isoGDGTs主要来源于浮游奇古菌,且有产甲烷古菌的碳源贡献。沉积物中5种isoGDGTs呈现显著的正相关关系(0.987 < R < 0.998,p < 0.01),表明各站的isoGDGTs具有一定的相同来源,其中GDGT-0与crenarchaeol之间有相关性,说明GDGT-0至少有产甲烷古菌和奇古菌的贡献[90]。生态系统的改变往往表现在微生物群落结构改变,而不同种类、不同功能群的古菌类群改变必然会影响RDOC的分子组成和生成速率及其MCP 储碳效率改变(增强或削弱)。科学研究发现,一些GDGTs的特定分子生物标志物就来自MCP活动,可确定生态环境中古菌类群结构和功能[91]。奇古菌作为古菌的突出代表,能够通过利用氧化氨根离子获取的能量进行CO2 的固定,在有机质降解代谢过程中改造并分泌RDOC[92],产生惰性的RDOC 储存于海洋碳库中。在碳循环中产甲烷古菌是极为重要的甲烷生产者,在海洋厌氧环境下,这类古菌可以将有机物降解为甲烷[93]。罗斯海表层沉积物中5种isoGDGT(GDGT-0、GDGT-1、GDGT-2、GDGT-3、GDGT-Crenarchaeol,)呈现显著的正相关关系(0.99 < R < 1,p < 0.01)。此外,isoGDGTs与brGDGTs含量呈现显著的线性关系(R = 0.94,p < 0.01,n = 15),且isoGDGTs和brGDGTs均与TOC含量呈现显著的线性关系(R = 0.69,p < 0.01;R = 0.64,p < 0.05,n = 15)有一致性分布特征,表明海洋古菌对表层沉积有机碳具有重要的贡献,且以上指标适用于罗斯海范围内产生isoGDGTs的古菌类群的重建。
4.1.3 利用表层沉积物
${\mathrm{TEX}}^{{\mathrm{L}}}_{\mathrm{86}}$ 重建罗斯海现代表层海水温度(SST)海表温度是影响浮游植物光合作用和固碳效率的重要因素[94]。2002年Schouten 等分析全球海区44个表层沉积物样品,首次提出基于GDGTs II、III、IV及V′的相对比值的TEX86指数(the Tetra Ether index of tetraethers consisting of 86 carbon atoms),并发现TEX86与年平均表层海水温度(SST)有很好的相关性(R = 0.92),建立了线性方程式[16],该方程的适用温度范围为0~30℃)
$$ {\mathrm{TEX}}_{86}= \frac{(\left[\mathrm{G}\mathrm{D}\mathrm{G}\mathrm{T}-2\right]+[\mathrm{G}\mathrm{D}\mathrm{G}\mathrm{T}-3]+[\mathrm{C}\mathrm{r}\mathrm{e}{\mathrm{n}}^{'}\left]\right)}{\left(\right[\mathrm{G}\mathrm{D}\mathrm{G}\mathrm{T}-1]+\left[\mathrm{G}\mathrm{D}\mathrm{G}\mathrm{T}-2\right]+\left[\mathrm{G}\mathrm{D}\mathrm{G}\mathrm{T}-3\right]+\left[\mathrm{C}\mathrm{r}\mathrm{e}{\mathrm{n}}^{'}\right])}, $$ (1) $$ \mathrm{TEX}_{\mathrm{86}}=0.015T+0.28\qquad(R=0.92,\ n=44). $$ (2) 在Schouten等提出TEX86指数后,海洋科学家在不同海洋区域与环境下应用和校正TEX86-SST计算公式[88, 95–97],2010年Kim等[21]提出了适于极地低温区的新校正
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 公式,适应于T < 15℃。前人研究显示$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标在南极半岛[98]、普里兹湾[99–100]、北冰洋[101]极地海洋重建SST方面展现出较大的应用潜力[102–103]。$$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}}=\mathrm{l}\mathrm{o}\mathrm{g}\left(\frac{\left[\mathrm{G}\mathrm{D}\mathrm{G}\mathrm{T}-2\right]}{\left[\mathrm{G}\mathrm{D}\mathrm{G}\mathrm{T}-1\right]+\left[\mathrm{G}\mathrm{D}\mathrm{G}\mathrm{T}-2\right]+\left[\mathrm{G}\mathrm{D}\mathrm{G}\mathrm{T}-3\right]}\right), $$ (3) $$ T=67.5\times\mathrm{T}\mathrm{E}\mathrm{X}_{86}^{\mathrm{L}}+46.9\quad(R=0.86\text{,}n=396\text{,}p < 0.000\; 1). $$ (4) 陆源有机质与海源有机质的参考比值BIT ( Branched and Isoprenoid Tetraether),BIT比值(GDGT Ia +GDGT IIa + GDGT IIIa)/Cren + GDGT Ia + GDGT IIa + GDGT IIIa) [104],一般认为在大洋区BIT值为0,表示海洋源有机质的贡献较高,而土壤的BIT值为1,表示陆源有机质的贡献最高。罗斯海表层沉积物中BIT值均较低0.02~0.11,平均为0.05(图6g),小于 BIT 阀值 0.4,表明沉积物中brGDGTs可能是海洋水体和沉积物中原位耐寒细菌所产生,而非陆源土壤的厌氧细菌输入,表明 TEX86 可以较真实地反映 SST 变化[105]。基于 isoGDGTs 与温度之间的线性关系建立罗斯海表层沉积物反演的 TEX86-SST 和
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 所计算的温度,与世界海洋图集及数据库(WOA2013,1955−2012 年数据集,分辨率为 0.25°,http://odv.awi.de/data/ocean/world-ocean-atlas-2013/ )中对应站位 0~200 m 海水各层温度,逐季逐月逐层与进行对比分析,发现TEX86-SST 公式(3)(T = 56.2 × TEX86 − 10.78)[95]和公式(4)( T = TEX86 −0.3038 /0.0125 )[96]所计算的温度TEX86-SST均偏高,而$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST公式(1)(T = 67.5 ×$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ + 46.9)[21]和公式(2)(T = 50.8 ×$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ + 36.1)[98] 所计算的各水层温度接近,整体上反演WOA 3 月表层水温(图7a),且$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST [公式(2)]重建的各水层海温与 WOA夏季末 3 月表层水温,二者呈现出良好的线性关系(R = 0.78,p < 0.01,n = 15)(图7b)。公式(2)比公式(1)更加适用于研究海区。罗斯海融冰期,形成开阔水域和冰间湖,初级生产力高,春季(11 月中旬至12 月)混合层接近 50 m,而夏季为 10 m 左右[100]。 图 7 罗斯海WOA 3月(a)和WOA夏季(b)表层温度与
图 7 罗斯海WOA 3月(a)和WOA夏季(b)表层温度与$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST的线性关系Figure 7. Linear relationship between Ross Sea WOA March (a) and WOA summer (b) surface temperature vs$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST4.1.4 藻类分子标志物与浮游动植物生产力/群落结构及其海洋BP作用
海洋生物泵(BP)过程的起始环节和关键部分是浮游植物群落,其通过自身代谢提供相当大的一部分DOC,浮游植物生物量决定了上层海洋初级生产力,其生产力愈强,吸收CO2就越多,故初级生产力可表征浮游植物固碳强度[106],初级生产力和生物碳泵(BCP)效率有紧密关联。目前3种藻类分子生物标志物(菜籽甾醇-brassicasterol、甲藻甾醇-dinosterol、C37长链烯酮-C37 alkenones)可重建浮游植物单一种群(硅藻、甲藻和颗石藻)生物量或群落结构变化[102],它们的总含量(SUM = 菜籽甾醇+甲藻甾醇 + C37长链烯酮)可直接指示浮游植物初级生产力变化[103, 107],群落结构变化/初级生产力在全球碳循环中起重要作用,其储碳效率在很大程度上决定了海洋和大气中的碳库变动[108–109]。硅藻是蛋白石骨骼,硅藻初级生产不仅可以将大气中的CO2固定为有机质,形成过程中只会吸收CO2,其硅质壳体还可以作为生物泵的“压舱物”加快上层有机质的沉降速率,以提高生物泵效率[110];而颗石藻是方解石骨骼,进行光合作用的同时发生钙化作用,在形成其碳酸钙骨骼壳体时,会向水体中释放出CO2,对生物泵而言是一种负面效应[3]。可见,浮游植物群落结构变化会影响着罗斯海生物泵效率[12, 111]。
罗斯海表层沉积物中3种藻类分子生物标志物总量SUM为0.36~275.86 ng/g(平均50.54 ng/g),菜籽甾醇-brassicasterol含量为0.14~198.41 ng/g(平均39.09 ng/g),甲藻甾醇-dinosterol含量为0.04~37.31 ng/g(平均6.00 ng/g),C37长链烯酮-C37 alkenones含量为0.18~40.12 ng/g(平均5.45 ng/g),菜籽甾醇-brassicasterol含量普遍占绝对优势(图8a)。罗斯海表层沉积物中菜籽甾醇-brassicasterol 和甲藻甾醇-dinosterol 占总量 SUM 的比例分别为 38.2%~86.88%(平均 71.01)、6.71%~20.25%(平均 8.69%),这与现场调查(夏季)浮游植物以硅藻为主的结果一致[112]。菜籽甾醇−硅藻与甲藻甾醇−甲藻的比值(bra/dino)为 2.16~12.73(平均 6.79),通常浮游植物群落中硅藻的占比与生物泵效率呈正相关关系,硅藻占比越高POC 输出通量越高,重建的硅藻与甲藻之间均有显著的相关性(R = 0.98,p < 0.01,n = 18),并与海冰消融的时空分布模式相一致,以上重建的浮游植物/初级生产力的空间分布,以及硅藻/甲藻占比优势均与实测生物数据相吻合[88],表明藻类分子标志物应用于重建罗斯海浮游植物初级生产力和优势种群具有很好的适用性。硅藻是罗斯海浮游植物的优势藻种,在每年夏季暴发性生长[58],即硅质泵占主导地位,而颗石藻因南极温度低不适宜生长,因此钙质泵对罗斯海的贡献是很小的。对比多年的生物实测数据,罗斯海浮游植物含量在海冰边缘及冰山周围丰度较高的结果相一致[66],表明冰架前缘沉积指示了高能的海洋环境,其分布特征与罗斯海冰间湖较高的初级生产力[56]和沉积物 TOC 含量的分布模式(图3a)相一致。南极春季首先吹开罗斯海南端和 Terra Nova Bay 的海冰而形成冰间湖,初级生产较高,尤其西南部冰间湖海水叶绿素 a 和初级生产均显著高于其他罗斯海区域,初级生产作为生物泵的起点,决定了浮游植物光合作用固定 CO2 的强弱[47],因此造成同一个海域具有明显不同的生物泵效率。
 图 8 现代表层沉积物的浮游动植物分子生物标志物含量平面分布a. 菜籽甾醇-brassicasterol(硅藻)含量,b. 甲藻甾醇−dinostero(甲藻)含量,c. 长链烯酮-long-chain alkenones(颗石藻)含量,d. 胆甾醇-Chlesterol(浮游动物总量)含量,e. SUM-初级生产力,f. 菜籽甾醇-硅藻占比(bra/sum),g. 硅藻/甲藻比值(bra/dino)Figure 8. Molecular biomarker contents of planktonic algae in modern surface sedimentsa. Brassasterol (diatoms) content, b. dinosterol (dinoflagellates) content, c. long-chain alkenones (ccolithophores) content, d. cholestanol (total zooplankton) content, e. SUM-primary productivity content, f. brassicasterol-silica ratio (bra/sum), g. diatom/dinoflagellate ratio (bra/dino)
图 8 现代表层沉积物的浮游动植物分子生物标志物含量平面分布a. 菜籽甾醇-brassicasterol(硅藻)含量,b. 甲藻甾醇−dinostero(甲藻)含量,c. 长链烯酮-long-chain alkenones(颗石藻)含量,d. 胆甾醇-Chlesterol(浮游动物总量)含量,e. SUM-初级生产力,f. 菜籽甾醇-硅藻占比(bra/sum),g. 硅藻/甲藻比值(bra/dino)Figure 8. Molecular biomarker contents of planktonic algae in modern surface sedimentsa. Brassasterol (diatoms) content, b. dinosterol (dinoflagellates) content, c. long-chain alkenones (ccolithophores) content, d. cholestanol (total zooplankton) content, e. SUM-primary productivity content, f. brassicasterol-silica ratio (bra/sum), g. diatom/dinoflagellate ratio (bra/dino)表层沉积物中胆甾醇-Chlesterol主要由各种海洋浮游动物产生及少量光合自养生物产生,可用于反演浮游动物总量的变化[113–114],浮游动物摄食和粪球输出是海洋生物泵的关键过程。利用胆甾醇-Chlesterol生物标志物重建的浮游动物总生产力为24.00~
3097.55 ng/g(平均548.51 ng/g),在维多利亚地陆架前缘和罗斯冰架前缘分别出现高值(R04站为3097.55 ng/g,JB01站1122.07 ng/g),表明浮游动物在罗斯海生态系统的能量流动中发挥重要作用。在南极磷虾是南极生态系生物量最大的浮游动物[115],故可反映磷虾总生产力变动,浮游甲壳类可通过直接分泌释放一定量的不稳定 DOC,其粪便溶解的 DOC 是水体 DOC 来源的另一途径[116],在南极磷虾拥有巨大生物量,因此磷虾生物泵效率不可忽视。此外,胆甾醇(南极磷虾)与菜籽甾醇(硅藻)和甲藻甾醇(甲藻)具有一致的分布特征(R = 0.88,R = 0.927,p < 0.01,n = 18)。浮游甲壳类也可以通过直接分泌释放一定量的不稳定DOC,其粪便溶解的DOC是水体DOC来源的另一途径[58],还有原生动物(最原始、最简单、最低等动物)对浮游植物摄食过程也同样伴随DOC的释放[88]。浮游植物生物标志物反演的各种藻类占比与
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标重建的SST-2具有一定相关性(图9a和b),菜籽甾醇比例与$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标重建的SST呈负相关关系,而甲藻甾醇呈现正相关关系,意味着在低温区喜冷的硅藻相对生物量较高,而在相对高温区,喜暖的甲藻相对生物量升高,这与现场调查硅藻在海冰边缘及冰山周围丰度较高的结果一致[58],在低温区域硅藻的占比高于甲藻,而在温度较高区域其含量较低。 图 9 硅藻和甲藻占比与重建表层海温SST的线性关性a. 菜籽甾醇/总量(bra/sum)和b. 菜籽甾醇/甲藻甾醇(bra/dino)与SST的相关性Figure 9. The linear correlation between the proportions of diatoms and dinoflagellates and the reconstructed SSTa. The correlation of brassicasterol/sum (bra/sum) and b. brassicasterol/dinosterol (bradino) with SST
图 9 硅藻和甲藻占比与重建表层海温SST的线性关性a. 菜籽甾醇/总量(bra/sum)和b. 菜籽甾醇/甲藻甾醇(bra/dino)与SST的相关性Figure 9. The linear correlation between the proportions of diatoms and dinoflagellates and the reconstructed SSTa. The correlation of brassicasterol/sum (bra/sum) and b. brassicasterol/dinosterol (bradino) with SST4.2 浮游生物生产力/结构变化与古海洋生物碳泵演化及其地质事件的联系
4.2.1 建立JB03岩芯年代框架及其沉积速率变化
利用10个层位中的全碳样品为材料,在Beta实验室完成AMS14C测试分析,将总有机碳测得AMS14C年龄扣除老碳污染(
3045 a)及碳库年龄(825 a),年代控制点之间和控制点之外采用Calb.7.10软件和Marine13程序(http://calib.org/calib/calib.html )进行日历年转换,年代控制点之间和控制点之外分别采用线性内插和线性外推方法来建立年代框架,为历史资料的解译提供准确的地层时代标尺。如表1所示,罗斯海JB03岩芯底部131 cm沉积物年龄为27.27 ka BP,岩芯131~95 cm分别对应于末次冰盛期(27.27~21.0 ka BP),岩芯95~73cm对应于末次冰消期(21.2~11.7 ka BP),岩芯73~0.0 cm对应于全新世(11.7~0.0 ka BP)。通过9个年龄控制点,利用插值法计算获得岩芯各层位的沉积速率,在79 cm沉积速率最低,而在末次冰盛期的111~131 cm出现较高的沉积速率为100~400 cm/ka(表1所示),沉积速率的不均一性反映沉积环境的差异性,这与黄梦雪等[117]在罗斯海Joides海槽JB06岩芯(该时期沉积速率较低为2~3 cm/ka)的结果存在显著差异,且在末次冰盛期31.5~27 ka BP期间,JB06岩芯的冰山指标和IRD(>250 μm)含量出现峰值,说明这一时期存在冰山搬运沉积物物输入,罗斯冰架出现崩塌消退。罗斯冰架与全球冰盖面积达到最大的时间基本接近(33~26.5 ka BP)[118–119]。此外,研究发现末次冰盛期,Joides海槽罗斯冰架触底线(Grounding Line)最北的位置可达74°S以南[119],而JB03岩芯位于75.8°S,170.7°E区域,因此可推测末次冰盛期该岩芯111 cm层位以深可能受罗斯冰架崩塌或触底引起扰动或再沉积等影响。表 1 南极罗斯海JB03柱样的AMS14C年龄及校正年龄与沉积速率Table 1. AMS14C ages and calibrated ages with sedimentation rates of JB03 core from Ross Sea of Antarctica深度/cm 14C年龄/a BP 化石碳年龄/a 海洋储层年龄/a 日历年龄/a BP 沉积速率/(cm·ka−1) 地质年代 0~2 4470 ± 303045 825 600 全新世 2~4 4774 ± 303045 825 904 6.58 18~20 6950 ± 303045 825 3080 7.35 54~56 12655 ± 303045 825 8785 6.31 68~70 14730 ± 303045 825 10860 6.75 72~74 15720 ± 303045 825 11850 4.04 末次冰消期 78~80 28950 ± 303045 825 25080 0.45 末次盛冰期 102~104 31010 ± 303045 825 27140 11.7 110~112 31090 ± 303045 825 27220 100 112~114 31095 ± 303045 825 27225 400 128~130 31140 ± 303045 825 27270 356 4.2.2 古海洋初级生产力/生物碳源对大尺度气候/沉积环境的响应
生物泵与气候变化的相互作用(影响、响应和反馈)已经历了漫长的地球历史进程[47]。罗斯海JB03岩芯全样(131~0 cm)年龄为27.27~0.0 ka BP(表1和图10)。CPI和OPE分别为 1.29~2.07(平均 1.63)和 1.14~1.80(平均 1.36),二者呈现出良好的线性关系(R = 0.67,p < 0.01,n = 66),具有一定的奇偶优势,说明该岩芯中长链饱和烃降解较少,有机质演化程度低,沉积有机质保存较好。自冰盛期、冰消期(受到冰架影响)至全新世暖期,古海洋初级生产力和生物碳源逐渐提高,全新世内源生物碳输入最高、冰盛期最低。
 图 10 AMS14C测年结果和校正后年龄及其沉积速率Figure 10. AMS14C dating results and calibrated ages and their depositional rates
图 10 AMS14C测年结果和校正后年龄及其沉积速率Figure 10. AMS14C dating results and calibrated ages and their depositional rates末次冰盛期(27.27~25.0 ka BP):表征古海洋初级生产力TOC(图11a)、海洋生物碳源的菌藻类生物量(nC15 + nC17 + nC19)和大型浮游植物生物量(nC21 + nC23 + nC25)(图11e和f)均低,显示出冰盛期生产者和初级生产力均低下,通过判断TOC/TN比值和正构烷烃分子特征,发现Pr/Ph和CPI指标(图11h和i)、陆源高等植被(nC27 + nC29 + nC31)和L/H(图11g和b)、海陆源比值TARHC(nC27 + nC29 + nC31)/(nC15 + nC17 + nC19)和(nC21 + nC22)/(nC28 + nC29)(图11c和d)等均呈现出剧烈的波动,暗示该地质历史时期存在剧烈的气候变化或地质演变,冰盛期沉积物除了来自表层生物碳源外,还有其他物质来源,如大量陆源有机物的输入(冰盛期风尘等陆源输入增加可能稀释了海源部分)。JB03岩芯(113~111 cm)层段的沉积速率异常高(100~400 cm/ka),该岩芯(111 cm层位以深)可能受到当时罗斯冰架崩塌或触底引起扰动,Joides海槽罗斯冰架触底线(Grounding Line)最北的位置可达74°S以南[117],而JB03岩芯位于75.8°S,170.7°E。25~21 ka BP期间受到再沉积的影响[117],故TOC以及低烃(nC15 + nC17 + nC19)和中烃(nC21 + nC23 + nC25)含量均低,说明当时海洋初级生产整体较低,沉积物储碳效率低下。
 图 11 JB03岩芯TOC/TN值和正构烷烃分子组合特征记录的末次冰期来古海洋陆源/海源碳库变化a. 总有机碳TOC和TOC/TN,b. 低碳烃/高碳烃L/H,c. TARHC,d. (nC21 + nC22)/(nC28 + nC29),e. 菌藻类(nC15 + nC17 + nC19),f. 大型浮游植物(nC21 + nC23 + nC25),g. 陆源高等植物(nC27 + nC29 + nC31),h. Pr/Ph,i. CPI,J. OEP值Figure 11. Last glacial marine/terrestrial carbon isotopic signature recorded by TOC/TN ratio and molecular composition n-alkanes from Core JB03a. TOC and TOC/TN, b. L/H, c. TARHC, d. (nC21 + nC22)/(nC28 + nC29) e. Fungi algae (nC15 + nC17 + nC19), f. Large planktonic plants (nC21 + nC23 + nC25), g. Terrestrial higher plants (nC27 + nC29 + nC31), h. Pr/Ph, i.CPI, j. OEP values
图 11 JB03岩芯TOC/TN值和正构烷烃分子组合特征记录的末次冰期来古海洋陆源/海源碳库变化a. 总有机碳TOC和TOC/TN,b. 低碳烃/高碳烃L/H,c. TARHC,d. (nC21 + nC22)/(nC28 + nC29),e. 菌藻类(nC15 + nC17 + nC19),f. 大型浮游植物(nC21 + nC23 + nC25),g. 陆源高等植物(nC27 + nC29 + nC31),h. Pr/Ph,i. CPI,J. OEP值Figure 11. Last glacial marine/terrestrial carbon isotopic signature recorded by TOC/TN ratio and molecular composition n-alkanes from Core JB03a. TOC and TOC/TN, b. L/H, c. TARHC, d. (nC21 + nC22)/(nC28 + nC29) e. Fungi algae (nC15 + nC17 + nC19), f. Large planktonic plants (nC21 + nC23 + nC25), g. Terrestrial higher plants (nC27 + nC29 + nC31), h. Pr/Ph, i.CPI, j. OEP values末次冰盛期到冰消期(21~11.8 ka BP前后):JB03岩芯记录的初级生产力TOC在0.7%左右波动(图11a),菌藻类(nC15 + nC17 + nC19)和大型浮游植物(nC21 + nC23 + nC25)生物量较低,表明该时期海洋生产者和初级生产力整体较低且稳定,意味着上层海洋BP效率不高。TOC/TN比值平均为13.16(图11),表明有大量陆源有机碳输入,同沉积地层中正构烷烃主峰碳主要为nC27,分子组合特征的L/H、(nC21 + nC22)/( nC28 + nC29)和TARHC值基本都小于1(图11),进一步佐证了这一时期,海洋来源的生物碳贡献小,沉积物有机质的富集可能受氧化还原条件与陆源冰伐碎屑输入或稀释了的海源有机质影响,因此埋藏效率低。前人研究发现,西南极冰心的WDC δ18O在22~21 ka BP前后开始偏重,气候变暖推动罗斯冰架开始缓慢消退(相较于罗斯冰架中部和东部),Joides海槽处冰架消退较早,在21 ka BP前后可能已退至74°S以南[117],罗斯海冰盖在15.2~13.9 ka BP前后开始全面后退[120]。岩芯记录的CPI和OEP值小,氧化还原指标(Pr/Ph)指示较强的氧化型沉积环境(图11),当富氧条件出现时,生物降解作用加强,这个沉积碳库可能会被氧化,不利于陆源和海洋生物有机碳的埋藏效率。
全新世(11.7~0.6 ka BP前后):岩芯记录的TOC含量平均为1.39(图11),明显高于末次冰期和冰消期,在10.27 ka BP前后出现一低值后迅速增加,可能是受到气候事件的影响。进入全新世后TOC/TN比值(图11)快速降低,而正构烷烃的主峰碳主要为低碳烃(nC17、nC18或nC19)、分子组合特征的L/H、(nC21 + nC22)/( nC28 + nC29)和TARHC(图11)则快速升高,基本大于1(除少数层位外),表明进入全新世以来,罗斯海冰筏碎屑输入减少,海洋生产者、初级生产力升高、海洋生物碳输入增加就意味着海洋BP效率加强,沉积有机质主要为海洋生物碳源。另外,进入全新世,沉积速率加快(6.27 cm/ka)(表1和图10),沉积环境的氧化性相对降低,有利于有机质的埋藏和保存。因此,全新世沉积有机碳(TOC平均1.39%)明显高于末次冰期和冰消期,也表明罗斯海古海洋初级生产力和生物碳源(增加/减少)对大尺度气候/环境演变是有显著响应的。
4.2.3 古海洋微生物/古菌类群与微生物碳泵演化对气候环境的响应
罗斯海JB03岩芯中微生物细胞膜脂(GDGTs)总量为216.53~
5634.78 ng/g(平均2519.98 ng/g),其中古菌细胞膜脂(isoGDGTs)含量为187.65~5277.58 ng/g(平均2327.13 ng/g),占GDGTs总量的87%~94%,isoGDGTs含量与TOC和SUM(浮游植物生物标志物总量)呈现出明显的相关性(分别为R = 0.81、R = 0.70、R = 0.87,p < 0.01,n = 40),这与表层沉积物结果相一致,进一步说明古细菌活动可能受到海洋初级生产力的影响,相对细菌更善于利用深海有机碳库中的DOC[121],从而产生更加惰性的RDOC于海洋碳库中。细菌细胞膜(brGDGTs)含量为28.88~357.20 ng/g(平均192.85 ng/g),占GDGTs的5.61%~13.34%。JB03岩芯中GDGTs、isoGDGTs、brGDGTs含量整体变化趋势与WDC δ18O、沉积物含水率具有一致的变化趋势(图12),GDGTs总量、isoGDGTs和brGDGTs含量与TOC均呈现显著的相关性(分别为R = 0.88、R = 0.88、R = 0.90,p < 0.01,n = 66),这与现代表层沉积物中GDGTs具有一致性结果。岩芯中古菌细胞膜脂(isoGDGTs)总量中以GDGT-0(产甲烷古菌+部分广古菌)和Crenarchaeol-(奇古菌)为主,GDGT-0在467.35~3003.12 ng/g范围,crenarchaeol在78.32~2122.26 ng/g范围,分别占isoGDGTs总量的51.92%~60.61%和36.15%~45.15%范围,不难发现产甲烷古菌和奇古菌是罗斯海古菌中主要类群。进一步根据R0/5值指示奇古菌和产甲烷古菌对古菌细胞脂isoGDGTs的贡献,岩芯沉积物中R0/5值为1.15~1.68(平均1.48),说明罗斯海JB03沉积物的isoGDGTs主要来源于浮游奇古菌,与表层沉积物结果相一致。 图 12 JB03岩芯微生物细胞膜脂GDGTs记录的古海洋微型生物/MCP地质演化a. WDC δ18O,b. 含水率和TOC,c. 微生物细胞膜脂-GDGTs,d. 古菌-IsoGDGTs,e. 细菌- brGDGTs,f. 产甲烷古菌- GDGT-0,g. 奇古菌-Crenarchaeol,h. R0/5比值,i. 菜籽甾醇- Brassicasterol(硅藻),j. 甲藻甾醇- Dinosterol(甲藻),k. C37长链烯酮- C37-ketene(颗石藻),l. 总量SUM-初级生产力,m. 胆甾醇-浮游动物总量Figure 12. Geological evolution of the microbial/MCP paleocean recorded by microbial cell membrane lipids GDGTs of JB03 corea. WDC δ18O, b. water content and TOC, c. microbial cell membrane lipids-GDGTs, d. archaeal-IsoGDGTs, e. bacterial- brGDGTs, f. Methanogen GDGT-0, g. Thaumarchaeota-crenarchaeol, h. R0/5 ratio, i. brassicol (diatoms), j. Dinosterol (dinoflagellates), k. C37 long-chain alken-C37-ketene (coccolithophores), l. Total SUM-primary production, m. Cholesterol-Total zoplankton
图 12 JB03岩芯微生物细胞膜脂GDGTs记录的古海洋微型生物/MCP地质演化a. WDC δ18O,b. 含水率和TOC,c. 微生物细胞膜脂-GDGTs,d. 古菌-IsoGDGTs,e. 细菌- brGDGTs,f. 产甲烷古菌- GDGT-0,g. 奇古菌-Crenarchaeol,h. R0/5比值,i. 菜籽甾醇- Brassicasterol(硅藻),j. 甲藻甾醇- Dinosterol(甲藻),k. C37长链烯酮- C37-ketene(颗石藻),l. 总量SUM-初级生产力,m. 胆甾醇-浮游动物总量Figure 12. Geological evolution of the microbial/MCP paleocean recorded by microbial cell membrane lipids GDGTs of JB03 corea. WDC δ18O, b. water content and TOC, c. microbial cell membrane lipids-GDGTs, d. archaeal-IsoGDGTs, e. bacterial- brGDGTs, f. Methanogen GDGT-0, g. Thaumarchaeota-crenarchaeol, h. R0/5 ratio, i. brassicol (diatoms), j. Dinosterol (dinoflagellates), k. C37 long-chain alken-C37-ketene (coccolithophores), l. Total SUM-primary production, m. Cholesterol-Total zoplankton末次冰盛期(27.27~21.0 ka BP),由于受到冰盖持续生长和海冰限制作用的影响,罗斯海上层古海洋初级生产力(TOC)低下,微生物总量(GDGTs)、古菌生物量(isoGDGTs)、细菌生物量(brGDGTs)、产甲烷古菌(部分广古菌)-(GDGT-0)和奇古菌(Crenarchaeol)生物量变化小;浮游植物生产力SUM、浮游动物总量胆甾醇、硅藻和甲藻生物量出现小幅度波动,在寒冷时段,海洋生产者、浮游植物生产力和浮游动物总量低,意味着BP效率和MCP效率一样被削弱。另外,BP是含碳化合物通过生物过程从海洋表面输送到深海的机制,沉积环境氧化性较高,沉积速率慢、不利于海洋BP储碳。
冰消期(21.0~11.7 ka BP),在 15.2~13.9 ka BP 左右罗斯海冰盖开始全面后退,可能是受到冰架消退、释放冰伐碎屑及营养物质,以及较暖的北部水向南移所致,硅藻生物量略有提高趋势(图12),还促进古海洋微生物类群生长,同期的微生物生产力、古菌和细菌生物量也呈现缓慢上升趋势,海洋微型生物是 MCP 储碳主要贡献者,意味着罗斯海 MCP 作用开始加强。另一方面,大量融冰致使水温很低,海洋初级生产力、甲藻和颗石藻生物量、浮游动物总量变化速度缓慢,表明海洋生物碳泵储碳效率低。
全新世(11.7~0.6 ka BP前后):全新世开始为11.5 ka BP时期,气候普遍转暖但具有明显的波动性。在岩芯11.7~10 ka BP期间,初级生产力-TOC、微生物总量-GDGTs、古菌-isoGDGTs、细菌-brGDGTs、浮游植物生产力(SUM)和硅藻生物量呈现小幅下降,这可能是受冰消期向全新世过渡时的YD冷事件影响,YD事件结束,地球快速过渡到温暖的早全新世(从10~8 ka BP开始),初级生产力-TOC(平均1.39)、微生物总量-GDGTs、古菌生物量-isoGDGTs和细菌生物量-brGDGTs整体上迅速升高,且远远高于末次冰期和冰消期,推测该时段岩芯的上层海洋浮游生物生长很可能已不再受到罗斯冰架的影响,同时温暖的气候条件、海冰消退形成开阔海域、有利于浮游植物群落生长、浮游动物总量快速增加,促进微生物生产力、古菌和细菌快速增加。浮游植物生产力-SUM与奇古菌-Crenarchaeol相关性较好(R = 0.80, n = 66),显示海洋厌氧氨氧化活动可能受到海洋初级生产力的影响,这将使得MCP大大加强。当海洋菌藻类、超微型生物或古菌占优势地位时,尽管海洋生物泵效应较弱,但浮游植物产生的活性溶解有机质进一步刺激了细菌活性,从而导致呼吸作用的加强、这将使得MCP效率加强。罗斯海全新世以来生态系统则以MCP为主导实现碳的捕获。同时沉积速率明显增大、Pr/Ph比值逐渐降低(为弱还原−弱氧化性环境),说明当时海水是缺氧分层性较好,高的生产力往往导致沉积物环境氧化程度降低,生物降解作用较弱,有利于生物碳泵作用的有机质埋藏和保存,导致RDOC库的积累。
4.2.4 古海洋浮游生物生产力/群落结构与生物碳泵演化对气候环境的响应
JB03 岩芯沉积记录的3种浮游植物生物标志物SUM总量(Brassicasterol + Dinosterol + C37-ketene)为0.76~77.33 ng/g(平均13.09 ng/g),表明初级生产力较低,意味海洋生物泵效率也相对低。沉积物中菜籽甾醇−硅藻、甲藻甾醇−甲藻和C37长链烯酮−颗石藻含量分别为0.45~62.53 ng/g(平均10.20 ng/g)、0.08~9.70 ng/g(平均1.21 ng/g)、0.24~5.09 ng/g(平均1.68 ng/g),表明末次冰盛期以来,硅藻是上层海洋浮游植物的优势种。JB03岩芯中菜籽甾醇/浮游植物总量(硅藻/SUM)占比为 46.86%~82.51%(平均72.17%);甲藻甾醇/浮游植物总量(甲藻/SUM)占比为 5.27%~14.9%(平均 8.90%);C37长链烯酮/浮游植物总量(颗石藻/SUM)占比为 5.16%~43.81%(平均18.93%);菜籽甾醇/甲藻甾醇比值为3.24~13.78,岩芯记录的各分子生物标志物所占相对比例,同样显示出硅藻是浮游植物的优势种群,与表层沉积物浮游植物生物标志物以指示硅藻的菜籽甾醇为主的结果一致。硅藻(硅质生物)是罗斯海浮游植物的绝对优势藻种,在每年夏季爆发性生长[58],因此罗斯海的生物泵主要是由“硅质泵”所驱动,沉积记录的浮游植物初级生产力和种群生物量从冰消期开始逐渐增加,特别是全新世暖期以来显著增加(图12)。3种浮游植物分子生物标志物均呈现显著的相关性(0.72 < R < 0.96,p < 0.01,n = 40),亦与TOC呈现较好相关性(0.40 < R < 0.63,p < 0.05,n = 40),表明浮游植物分子生物标志物总量(SUM)可用于指示罗斯海初级生产力的变化。
胆甾醇含量为42.16~683.27 ng/g(平均154.37 ng/g),且与3种浮游植物分子生物标志物(菜子甾醇、甲藻甾醇、C37长链烯酮)和总量SUM均呈现显著的线性关系(R = 0.93,R = 0.93,R = 0.82 和 R = 0.93),表明随上层海洋初级生产力的增加,浮游动物的生物量亦增加,相比较于末次冰期及冰消期,全新世以来罗斯海浮游动物总量快速增加。值得关注的是,进入全新世开始,JB03 岩芯的胆甾醇占比急剧增大,意味着罗斯海南极浮游动物或磷虾生物量和总生产力提高,这种情况与南极半岛附近海域相似[122],浮游动物摄食和粪球输出是海洋生物泵的关键过程[123],粪球可供细菌和其他浮游微生物摄食并释放DOC[124],之后异养微生物利用DOC转化为RDOC[125]。南极磷虾其粪便具有很高的沉降速率,达 309 m/d(打包效应 Packing effect)[126],能够阻碍微生物的降解、加速沉降保存在海洋中,达到海洋生物碳泵(BCP)储碳的效果。因此南极磷虾在 DOC 释放、RDOC 碳源贡献及其南大洋生物碳泵中扮演重要角色。
4.2.5 全新世以来浮游植物占比/群落结构与
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ –SST变化/冷事件联系JB03岩芯使用TEX86和
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 所计算的SST-1和SST-2海表温度分别为−2.38~3.03℃(平均0.29℃)、−0.98~3.08℃(平均1.02℃),两者具有一致变化趋势(R = 1.0,p < 66,n = 66),如图13所示,TEX86重建的古海洋SST普遍高于现实温度,而$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标重建的SST,公式(2)(T = 50.8×$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ + 36.1)计算的结果相比于公式(1)(T = 67.5 ×$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ + 46.9)所反演的SST波动较小、更接近古海洋现实温度,且JB03岩芯$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标所计算的SST-2与菜籽甾醇−硅藻生物量和硅藻/总量SUM的占比均呈现出一定的负相关关系;而甲藻/SUM的占比和颗石藻/SUM的占比均与$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标所计算的SST-2呈现一定的正相关关系(图14),说明$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 公式(2)更适合罗斯海古海洋的重建。 图 13 JB03沉积地层
图 13 JB03沉积地层$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标计算的SST-1和SST-2Figure 13. Calculation of JB03 sedimentary stratigraphic index SST-1 and SST-2 图 14 分子生物标志重建的浮游植物相对生物量与SST-2和的相关性a. 菜籽甾醇−硅藻含量(ng/g)与SST-2(℃),b. brassicasterol/Sum(%)与SST-2(℃),c. dinosterol/Sum(%)与SST-2(℃),d. alkenone/Sum(%)与SST-2(℃)的相关性Figure 14. Correlation between the relative biomass of phytoplankton with molecular biomarker reconstruction and SST-2a. Brassicasterol-siliceous content (ng/g) with SST-2 (℃), b. brassicasterol/Sum (%) with SST-2 (℃), c. dinosterol/Sum (%) with SST-2 (℃), d. alkenone/Sum (%) with SST-2 (℃)
图 14 分子生物标志重建的浮游植物相对生物量与SST-2和的相关性a. 菜籽甾醇−硅藻含量(ng/g)与SST-2(℃),b. brassicasterol/Sum(%)与SST-2(℃),c. dinosterol/Sum(%)与SST-2(℃),d. alkenone/Sum(%)与SST-2(℃)的相关性Figure 14. Correlation between the relative biomass of phytoplankton with molecular biomarker reconstruction and SST-2a. Brassicasterol-siliceous content (ng/g) with SST-2 (℃), b. brassicasterol/Sum (%) with SST-2 (℃), c. dinosterol/Sum (%) with SST-2 (℃), d. alkenone/Sum (%) with SST-2 (℃)$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 重建的SST(图15)显示JB03岩芯在冰消期向全新世过渡时期,出现一个由暖期−冷期(11.36~10.27 ka BP)快速变化的气候事件,很可能记录的是晚冰期(Late Glacial)的波林阿路德暖期(Bolling-Allerod)−新仙女木期(Younger Dryas,YD)的气候快速变化事件,随后温度又快速升高。JB03岩芯地层(64~66 cm)对应于10.27 ka BP前后,$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST重建的古海洋表层温度降到最低(SST为−0.29℃),罗斯海受到YD冷事件影响海冰面积扩大,初级生产力(藻类指标总SUM)、浮游动植物(胆甾醇-浮游动物、菜籽甾醇-硅藻、甲藻甾醇−甲藻)、海洋古菌(主要是Crenarchaeol-奇古菌)和原位细菌(brGDGTs)的丰度均一致降低,这与全球气候发生快速变冷的YD事件在11~10 ka BP的时间恰好吻合[127],与吴时国和陆钧[128]在南极普里兹湾利用喜暖硅藻种(主要为Thalassiosira)与喜冷硅藻种(主要为Nitzschia curta和Fragilariopsis antarctica)绝对数量变化和优势种数量的变化记录了晚冰期波林阿路德暖期向YD转变的结果一致。YD冷事件后,罗斯海迅速升温进入全新世暖期,冰架继续消退,CDW上涌区域南移,进一步促进冰架的消退,这与全新世早期气候最暖期(Antarctic early Holocene Optimum,AHO)有密切关系[129–130]。在11.85~10.86 ka BP(对应于68~70 cm地层)呈现高值,$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST对应2.56℃,表明罗斯海古海洋受到全新世早期AHO气候极暖事件的影响,这在南大洋冰芯和大西洋扇区的沉积物、罗斯海ANT31−JB06岩芯的冰山指标和浮冰碎屑(IRD)含量都记录了AHO气候极暖事件[131]。硅藻相对比例和bra/dino的比值均呈现一个低谷,而甲藻和颗石藻相对比例则出现一个峰值,表明喜冷性硅藻相对生物量降低,而喜暖性颗石藻和甲藻的相对生物量增加,可能是对全新世南极气候最暖期的响应。 图 15 全新世以来浮游植物相对生物量和SST-2变化对重要地质冷事件的响应a. 硅藻-bra/浮游植物总量(%),b. 甲藻-dino/浮游植物总量(%),c. 颗石藻-alk/浮游植物总量(%),d. 硅藻-bra/甲藻-dino,e. $ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $-SST-2Figure 15. Relative biomass and SST-2 change response to important geological cold events since the Holocenea. Diatoms-bra/total phytoplankton (%), b. dinoflagellates-dino/total phytoplankton (%), c. coccolithophores-alk/total phytoplankton (%), d. diatoms-bra/oflagellates-dino, e. $ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $-SST-2
图 15 全新世以来浮游植物相对生物量和SST-2变化对重要地质冷事件的响应a. 硅藻-bra/浮游植物总量(%),b. 甲藻-dino/浮游植物总量(%),c. 颗石藻-alk/浮游植物总量(%),d. 硅藻-bra/甲藻-dino,e. $ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $-SST-2Figure 15. Relative biomass and SST-2 change response to important geological cold events since the Holocenea. Diatoms-bra/total phytoplankton (%), b. dinoflagellates-dino/total phytoplankton (%), c. coccolithophores-alk/total phytoplankton (%), d. diatoms-bra/oflagellates-dino, e. $ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $-SST-2然而,温暖的全新世气候也同样具有不稳定性,表现为多次冷事件的发生,根据Bond等[132]的研究,全新世共发生9次冷事件。JB03岩芯地层的
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标重建的海表温度SST-2以及浮游植物群落结构变化(图15),记录了全新世暖期罗斯海主要发生7次气候变冷事件,由于该岩芯沉积速率较低(表1和图10),分辨率不够高、不同的替代指标会有一定的时间偏差,但这些气候变冷事件与北大西洋浮冰碎屑(IRD)记录全新世气候变冷事件的时间相近。JB03 岩芯 58~60 cm层段和 52~54 cm 分别对应于 9.38 ka BP 和 8.47 ka BP 左右,$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 重建的表层海温 SST 分别下降至 1.59℃和 1.31℃,与古里雅冰芯记录揭示出全新世早期存在两次低温事件(9.4 ka BP 和 8.2 ka BP)相对应[133]。JB03 岩芯 44~40 cm 地层对应 6.57 ka BP前后,与全新世南极在 6.5 ka BP 前后开始变冷的时间相近[134],$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 重建的表层海温为−0.69℃和−0.3℃;38~40 cm 地层$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 重建的表层海温为−0.41℃,与 6.2 ka BP 全新世以来最冷的一个降温事件相吻合[134]。4.2 ka BP 是晚全新世最强一次气候突变降温事件[134],$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 重建的海表温度 SST-2 为−0.03℃;岩芯 22~24 cm地层对应年代 4.35 ka BP 前后,$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 重建的 SST-2 为−0.41℃,指示气候寒冷。全新世这4个冷时段(9.38 ka BP、8.2 ka BP前后 、6.57 ka BP前后、4.35 ka BP前后)的硅藻相对生物量和 bra/dino 比值均出现升高小峰值,即在这4个时期喜冷硅藻相对生物量增加,而甲藻和颗石藻的相对生物量均出现小小降低,即甲藻和颗石藻相对生物量降低,反映了气候变冷,海洋硅质泵效率增强、而钙质泵效率降低,与$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标重建 SST 记录气候变冷的结果一致,也说明浮游植物群落结构变化与初级生产力对全球性大尺度气候变化的响应有差异。在 JB03 岩芯(地层 16~18 cm)
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 重建的古海洋温度(SST-2 为 2.86℃),出现高值之后,在14~16 cm 对应于岩芯年代 2.81 ka BP ,气候呈现不可逆波动降温 SST-2 为−0.37℃,表明罗斯海全新世暖期的结束,与敦德冰芯 δ18O 记录的气候变化基本一致,与北大西洋(北欧和格陵兰)全新世暖期结束时间接近[135–136];在岩芯 10~12 cm 层段对应于年代 1.45 ka BP 前后,$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST 记录了一次 1.4 ka BP 冷事件,重建古海洋温度 SST-2 为−0.22℃,与北大西洋浮冰碎屑 IRD 记录的 1.4 ka BP冷事件的时间相近[134]。罗斯海在 2.81 ka BP 和 1.45 ka BP 前后发生的两次变冷事件,$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标重建的古海洋 SST 均呈现降低,在这 2 个冷事件期间硅藻相对生物量和 bra/dino 比值开始逐渐增加,即在这两个时期喜冷硅藻相对生物量增加;而甲藻和颗石藻相对生物量逐渐降低,即甲藻和颗石藻相对生物量降低。全新世末小冰期(0.5~0.1 ka BP),是距离我们最近的一次大规模降温事件,其寒冷程度超过了晚全新世的 2.8 ka BP 和 4.2 ka BP 事件[137],在 JB03 岩芯地层(0~2 cm)在 0.6 ka BP 前后,$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST 记录了南极罗斯海古海洋显著降温至−0.74℃。一些气候学家根据当前地球气候变化多端的趋势进行预测,担心小冰期再次来临,因此加强生物碳泵地质演化的研究[138]。5. 结论
(1) 本文利用表层沉积物类脂生物标志物反演的现代罗斯海沉积碳库来源,以海洋自身源为罗斯海最主要的海洋生物碳库,沉积物中菜籽甾醇(硅藻)和甲藻甾醇(甲藻)占总量 SUM 的比例平均 71.01%和8.69%,与现场调查(夏季)浮游植物以硅藻为主的结果一致,即硅质泵占主导地位,而颗石藻因南极温度低不适宜生长,因此钙质泵对罗斯海的贡献是很小的。另外沉积物胆甾醇重建的浮游动物总生产力较高,南极磷虾在南大洋拥有巨大生物量,因此磷虾生物泵效率不可忽视。
(2) 表层沉积物中5种isoGDGT呈现显著的正相关关系,与TOC含量呈现显著的线性关系,有一致性分布特征,表明海洋古菌对表层沉积有机碳具有重要的贡献。基于 isoGDGTs 与温度之间的线性关系建立罗斯海表层沉积物反演的 TEX86-SST 和
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 所计算的温度,进行对比研究发现$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST[ 公式(2)]比公式(1)更加适用于研究海区。(3) 反演的现代海洋各种藻类占比与
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标重建的SST-2具有一定相关性,菜籽甾醇比例与$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标重建的SST呈负相关关系,而甲藻甾醇呈现正相关关系,意味着在低温区喜冷的硅藻相对生物量较高,而在相对高温区,喜暖的甲藻相对生物量升高,这与现场调查硅藻在海冰边缘及冰山周围丰度较高的结果一致。(4) 末次盛冰期受冰盖和海冰限制作用,古海洋初级生产低,沉积速率低下,沉积物处在弱氧化−氧化的沉积环境,不利于MCP和BCP储碳,在寒冷时段海洋碳储量潜力低下。冰消期气候开始变暖、推动罗斯冰架后退,硅藻生物量开始略有提高趋势(硅质泵加强),也增强了微生物的活性,促进古菌和细菌等生长,意味着MCP作用有所加强。进入全新世暖期,温暖的气候有利于浮游植物初级生产力、浮游动物总量增加,进而影响有机碳向深海的输送,加强了BP运转效率,沉积速率明显增大,环境氧化性减弱,有利于BP生物碳的储存;期间细菌/古菌的生命活动随温度升高而增强,包括微生物死亡,其核心脂质(是海洋有机沉积物的显著成分)可能被纳入大颗粒(产生RDOC),迅速输入深海,埋入海洋沉积物里,即碳储量增加,全新世的海洋碳循环与全球气候变暖密切相关。
(5) 全新世以来,
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标重建的海表温度SST与浮游植物占比(相对生物量)/群落结构变化,记录了全新世暖期罗斯海主要发生了7次气候变冷事件,在这冷事件期间硅藻相对生物量和bra/dino比值开始逐渐增加,即喜冷硅藻相对生物量增加;而甲藻和颗石藻相对生物量逐渐降低,即甲藻和颗石藻相对生物量降低,说明罗斯海浮游植物群落结构变化与全球大尺度气候变化具有密切关系。致谢:衷心感谢参加中国南极科学考察任务的全体科考人员和“雪龙”号上的全体船员,感谢南北极环境综合考察与评估专项的联合资助,感谢二位审稿人对论文提出了许多宝贵意见,在此一并致谢。
-
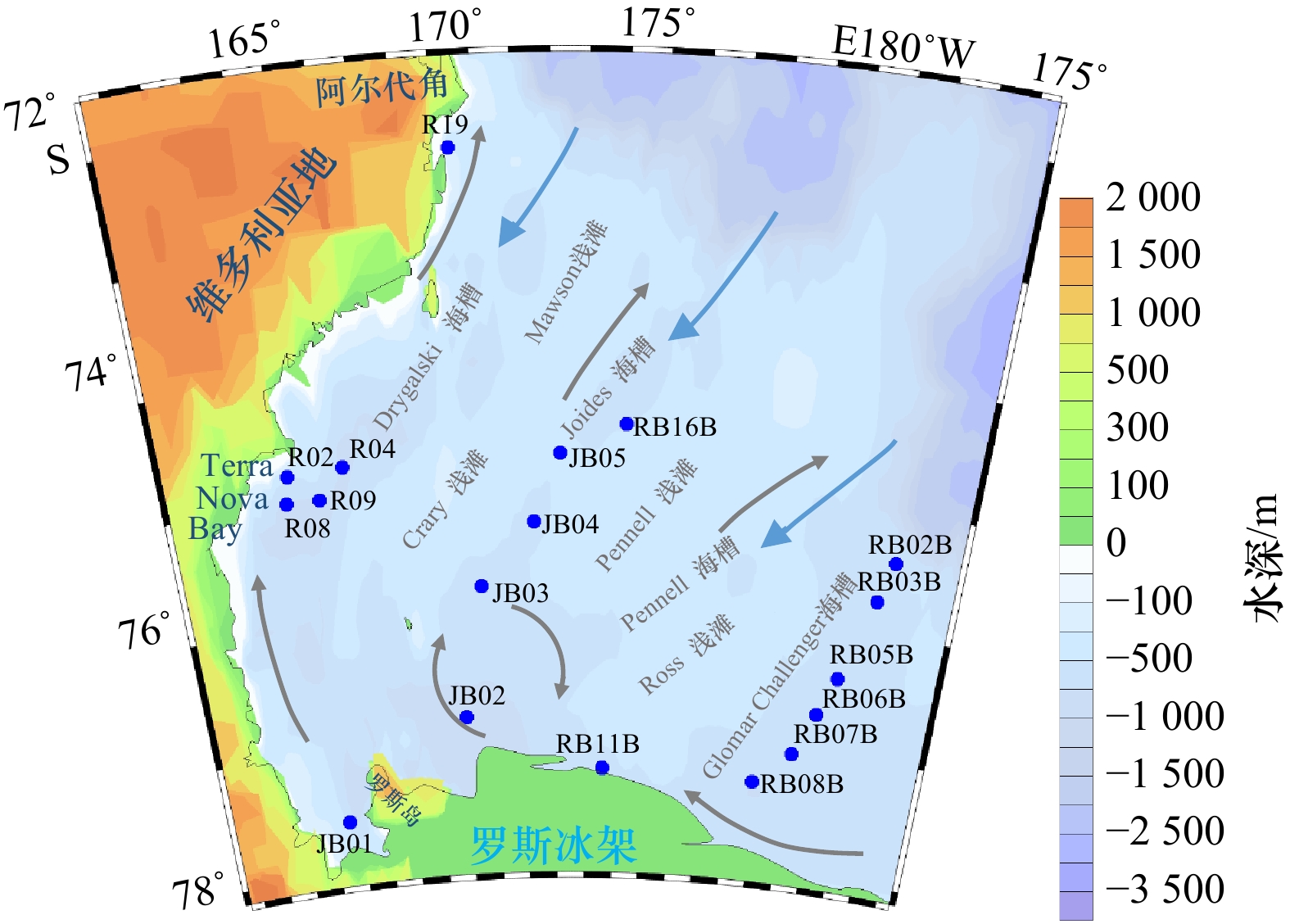
图 1 罗斯海表层沉积物和岩芯采样站位
箭头代表流向,灰色为表层环流,蓝色为变性绕极深层水(MCDW)涌入陆架
Fig. 1 Ross Sea surface sediment and core sampling stations
Arrows represent flow direction, grey represent the surface gyre, and blue represent the Modified Circumpolar Deep Water (MCDW) spilling onto the shelf

图 2 表层沉积物(a)正构烷烃(RB08B、JB03和RB06B)和(b)甾醇(JB01)气相及(c)GDGTs(JB01)液相色谱图
Fig. 2 Gas (a) and liquid (c) chromatograms of the surface sediments for (a) n-alkanes (RB08B, JB03, and RB06B) and (b) sterols (JB01) and (c) GDGTs (JB01)

图 3 a. 初级生产力-TOC含量(%),b. 总有机氮-TN含量(%),c. T/N比值
Fig. 3 a. Primary productivity-TOC content (%), b. total organic nitrogen-TN content (%), c. T/N ratio
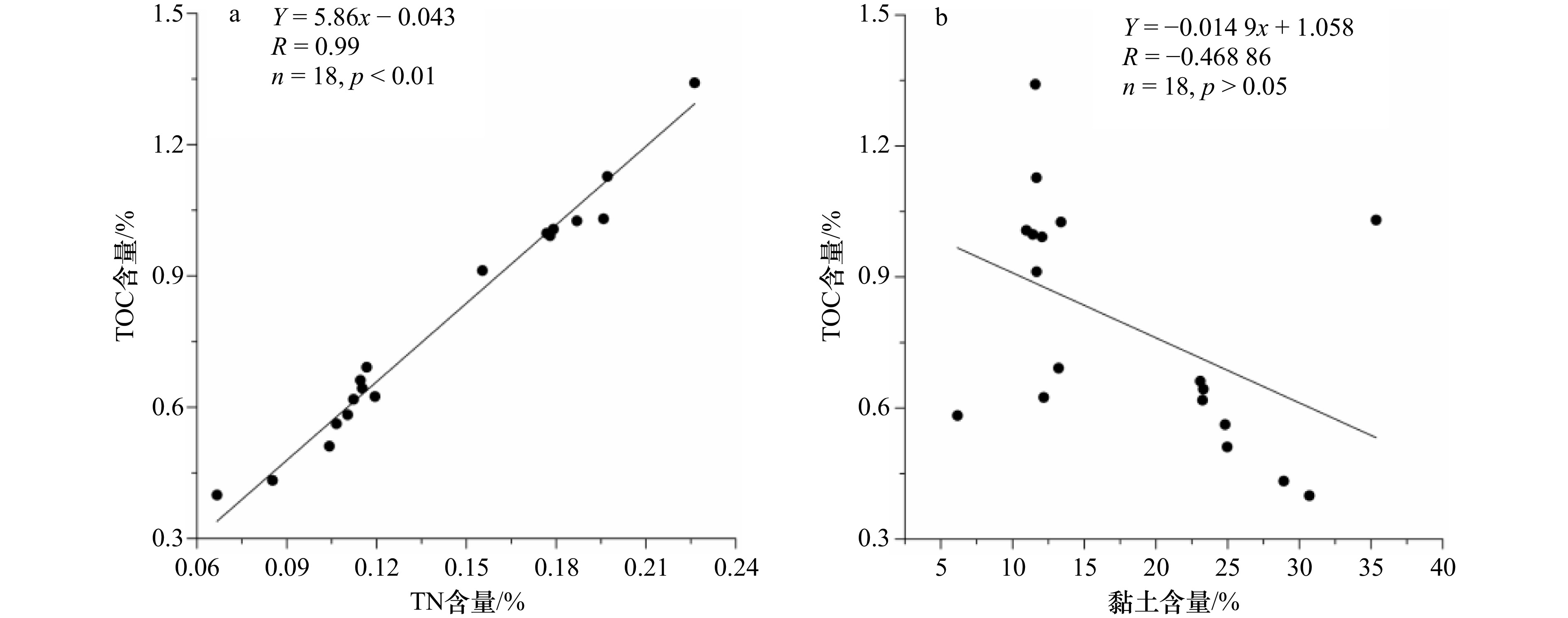
图 4 a. TOC含量(%)与TN含量(%),b. TOC含量(%)与黏土含量相关性
Fig. 4 a. TOC content (%) vs TN content (%), b. TOC content (%) vs clay content correlation

图 5 现代表层沉积物中TOC和正构烷烃总量及其分子组合特征与海洋生物泵联系
a. 正构烷烃总量(∑n-ALK),b. 菌藻类(nC15+nC17+nC19),c. 大型浮游植物(nC21+nC23+nC25),d. 陆源高等植物(nC27+nC29+nC31),e. 姥鲛烷/植烷(Pr/Ph),f. Ph/nC17值,g. Pr/nC18值,h. L/H值,i. (nC21+nC22)/(nC28+nC29)值,j. CPI值 ,k. OEP值
Fig. 5 Comparison of TOC and total normal alkane and its molecular composition characteristics in modern sediments with marine biological pump
a. Normal alkane (∑n-ALK) , b. fungi algae (nC15+ nC17 +nC19), c. large phytoplankton (nC21+nC23+nC25), d. terrestrial higher plants (nC27+nC29+nC31), e. cetane/phytane (Pr/Ph), f. Ph/nC17, g. Pr/nC18, h. L/H, i. (nC21+nC22)/(nC28+nC29), j. CPI, k. OEP

图 6 现代表层沉积物微生物细胞膜脂GDGTs、不同类群的古菌及其与MCP储碳联系
a. 微生物细胞膜脂-GDGTs含量,b. 古菌-IsoGDGTs含量,c. 细菌-BrGDGTs含量,d. 产甲烷古菌-GDGT-0含量,e. 奇古菌-Crenarchaeol含量,f. R0/5值,g. BIT值
Fig. 6 Modern surface sedimentary microbial cell membrane GDGTs, archaeal groups and their link to MCP carbon storage
a. Microbial cell membrane lipids-GDGTs content, b. Archaea-IsoGDGTs content, c. Bacteria-BrGDGTs content, d. Methanogenic archaea-GDGT-0 content, e. Thaumarchaeota-Crenarchaeol content, f. R0/5 values, g. BIT values

图 7 罗斯海WOA 3月(a)和WOA夏季(b)表层温度与
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST的线性关系Fig. 7 Linear relationship between Ross Sea WOA March (a) and WOA summer (b) surface temperature vs
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ -SST
图 8 现代表层沉积物的浮游动植物分子生物标志物含量平面分布
a. 菜籽甾醇-brassicasterol(硅藻)含量,b. 甲藻甾醇−dinostero(甲藻)含量,c. 长链烯酮-long-chain alkenones(颗石藻)含量,d. 胆甾醇-Chlesterol(浮游动物总量)含量,e. SUM-初级生产力,f. 菜籽甾醇-硅藻占比(bra/sum),g. 硅藻/甲藻比值(bra/dino)
Fig. 8 Molecular biomarker contents of planktonic algae in modern surface sediments
a. Brassasterol (diatoms) content, b. dinosterol (dinoflagellates) content, c. long-chain alkenones (ccolithophores) content, d. cholestanol (total zooplankton) content, e. SUM-primary productivity content, f. brassicasterol-silica ratio (bra/sum), g. diatom/dinoflagellate ratio (bra/dino)
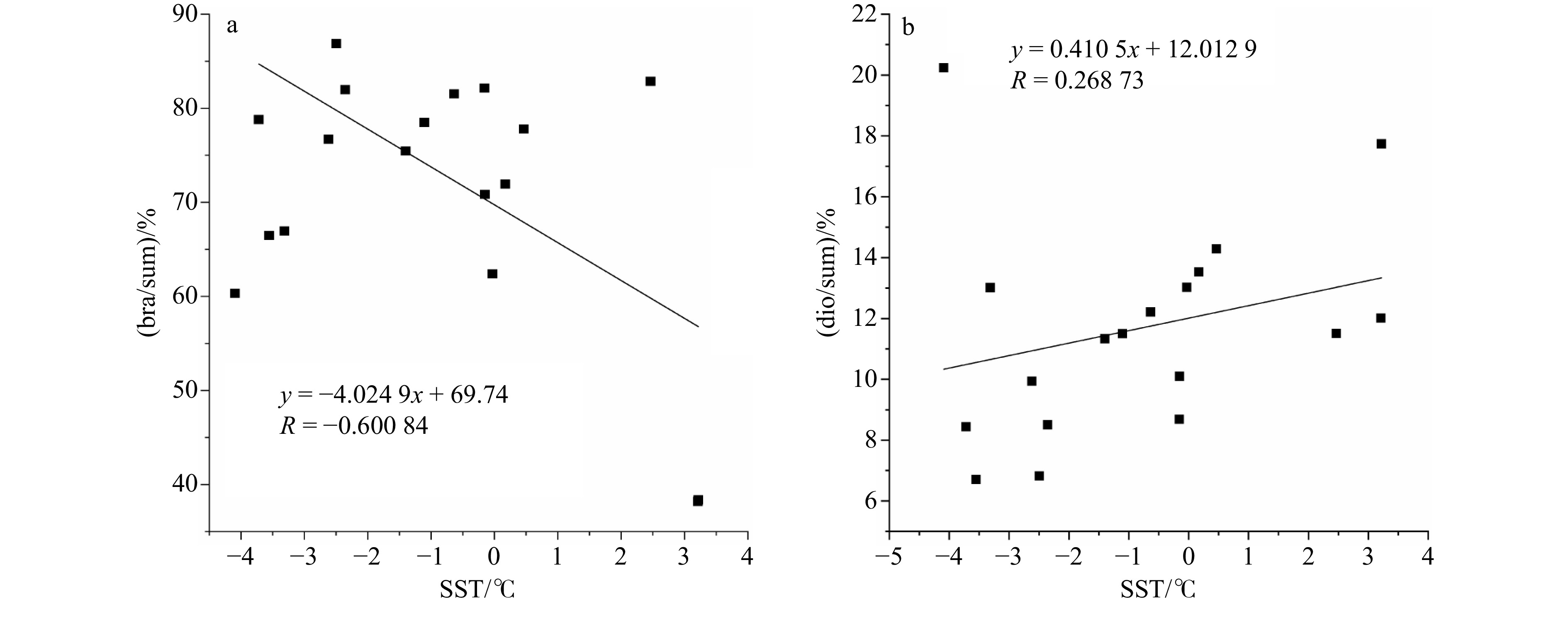
图 9 硅藻和甲藻占比与重建表层海温SST的线性关性
a. 菜籽甾醇/总量(bra/sum)和b. 菜籽甾醇/甲藻甾醇(bra/dino)与SST的相关性
Fig. 9 The linear correlation between the proportions of diatoms and dinoflagellates and the reconstructed SST
a. The correlation of brassicasterol/sum (bra/sum) and b. brassicasterol/dinosterol (bradino) with SST

图 10 AMS14C测年结果和校正后年龄及其沉积速率
Fig. 10 AMS14C dating results and calibrated ages and their depositional rates

图 11 JB03岩芯TOC/TN值和正构烷烃分子组合特征记录的末次冰期来古海洋陆源/海源碳库变化
a. 总有机碳TOC和TOC/TN,b. 低碳烃/高碳烃L/H,c. TARHC,d. (nC21 + nC22)/(nC28 + nC29),e. 菌藻类(nC15 + nC17 + nC19),f. 大型浮游植物(nC21 + nC23 + nC25),g. 陆源高等植物(nC27 + nC29 + nC31),h. Pr/Ph,i. CPI,J. OEP值
Fig. 11 Last glacial marine/terrestrial carbon isotopic signature recorded by TOC/TN ratio and molecular composition n-alkanes from Core JB03
a. TOC and TOC/TN, b. L/H, c. TARHC, d. (nC21 + nC22)/(nC28 + nC29) e. Fungi algae (nC15 + nC17 + nC19), f. Large planktonic plants (nC21 + nC23 + nC25), g. Terrestrial higher plants (nC27 + nC29 + nC31), h. Pr/Ph, i.CPI, j. OEP values
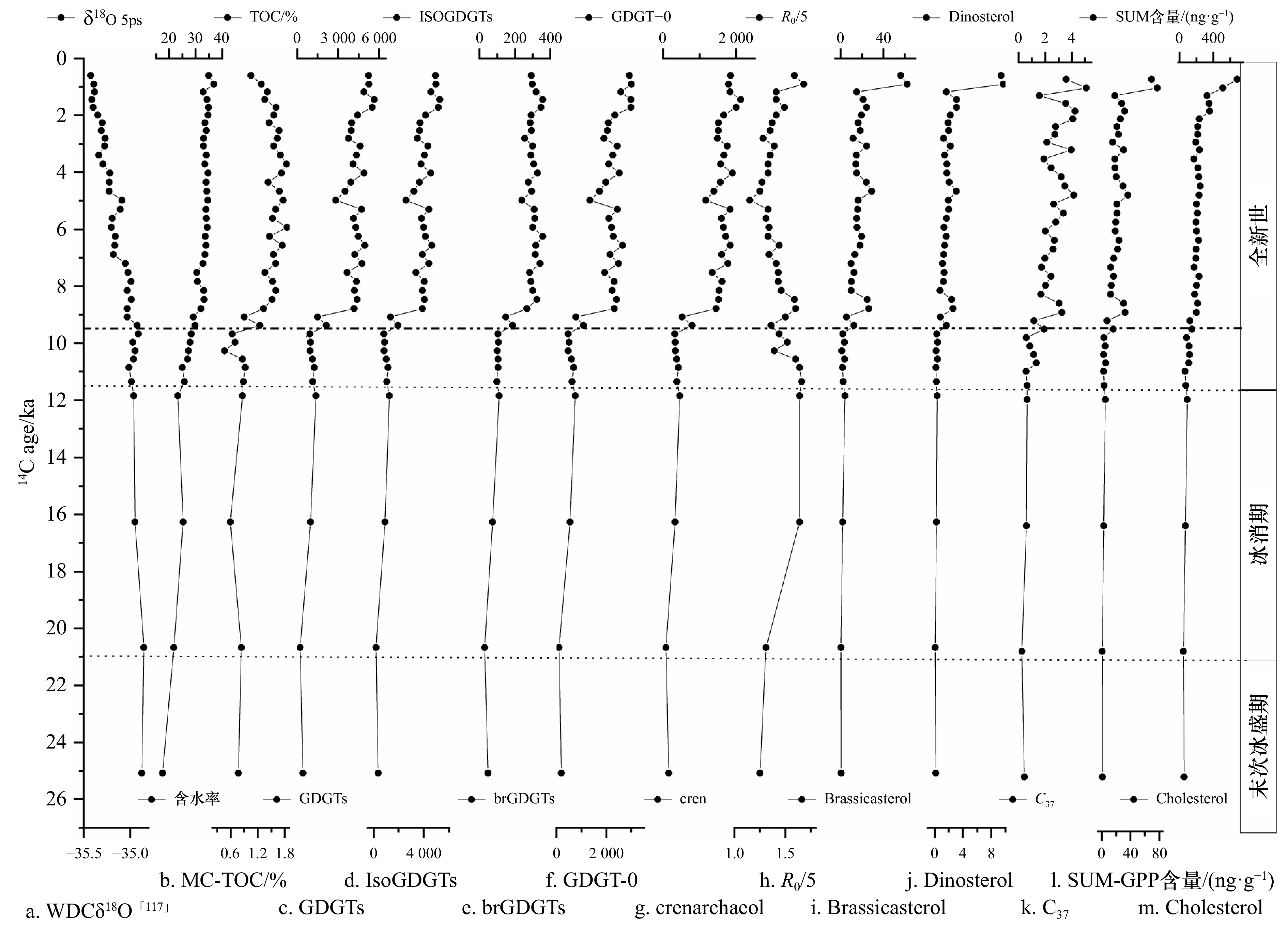
图 12 JB03岩芯微生物细胞膜脂GDGTs记录的古海洋微型生物/MCP地质演化
a. WDC δ18O,b. 含水率和TOC,c. 微生物细胞膜脂-GDGTs,d. 古菌-IsoGDGTs,e. 细菌- brGDGTs,f. 产甲烷古菌- GDGT-0,g. 奇古菌-Crenarchaeol,h. R0/5比值,i. 菜籽甾醇- Brassicasterol(硅藻),j. 甲藻甾醇- Dinosterol(甲藻),k. C37长链烯酮- C37-ketene(颗石藻),l. 总量SUM-初级生产力,m. 胆甾醇-浮游动物总量
Fig. 12 Geological evolution of the microbial/MCP paleocean recorded by microbial cell membrane lipids GDGTs of JB03 core
a. WDC δ18O, b. water content and TOC, c. microbial cell membrane lipids-GDGTs, d. archaeal-IsoGDGTs, e. bacterial- brGDGTs, f. Methanogen GDGT-0, g. Thaumarchaeota-crenarchaeol, h. R0/5 ratio, i. brassicol (diatoms), j. Dinosterol (dinoflagellates), k. C37 long-chain alken-C37-ketene (coccolithophores), l. Total SUM-primary production, m. Cholesterol-Total zoplankton

图 13 JB03沉积地层
$ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $ 指标计算的SST-1和SST-2Fig. 13 Calculation of JB03 sedimentary stratigraphic index SST-1 and SST-2

图 14 分子生物标志重建的浮游植物相对生物量与SST-2和的相关性
a. 菜籽甾醇−硅藻含量(ng/g)与SST-2(℃),b. brassicasterol/Sum(%)与SST-2(℃),c. dinosterol/Sum(%)与SST-2(℃),d. alkenone/Sum(%)与SST-2(℃)的相关性
Fig. 14 Correlation between the relative biomass of phytoplankton with molecular biomarker reconstruction and SST-2
a. Brassicasterol-siliceous content (ng/g) with SST-2 (℃), b. brassicasterol/Sum (%) with SST-2 (℃), c. dinosterol/Sum (%) with SST-2 (℃), d. alkenone/Sum (%) with SST-2 (℃)

图 15 全新世以来浮游植物相对生物量和SST-2变化对重要地质冷事件的响应
a. 硅藻-bra/浮游植物总量(%),b. 甲藻-dino/浮游植物总量(%),c. 颗石藻-alk/浮游植物总量(%),d. 硅藻-bra/甲藻-dino,e. $ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $-SST-2
Fig. 15 Relative biomass and SST-2 change response to important geological cold events since the Holocene
a. Diatoms-bra/total phytoplankton (%), b. dinoflagellates-dino/total phytoplankton (%), c. coccolithophores-alk/total phytoplankton (%), d. diatoms-bra/oflagellates-dino, e. $ {\mathrm{T}\mathrm{E}\mathrm{X}}_{86}^{\mathrm{L}} $-SST-2
表 1 南极罗斯海JB03柱样的AMS14C年龄及校正年龄与沉积速率
Tab. 1 AMS14C ages and calibrated ages with sedimentation rates of JB03 core from Ross Sea of Antarctica
深度/cm 14C年龄/a BP 化石碳年龄/a 海洋储层年龄/a 日历年龄/a BP 沉积速率/(cm·ka−1) 地质年代 0~2 4470 ± 303045 825 600 全新世 2~4 4774 ± 303045 825 904 6.58 18~20 6950 ± 303045 825 3080 7.35 54~56 12655 ± 303045 825 8785 6.31 68~70 14730 ± 303045 825 10860 6.75 72~74 15720 ± 303045 825 11850 4.04 末次冰消期 78~80 28950 ± 303045 825 25080 0.45 末次盛冰期 102~104 31010 ± 303045 825 27140 11.7 110~112 31090 ± 303045 825 27220 100 112~114 31095 ± 303045 825 27225 400 128~130 31140 ± 303045 825 27270 356  下载: 导出CSV
下载: 导出CSV
-
[1] 谢树成, 焦念志, 罗根明, 等. 海洋生物碳泵的地质演化: 微生物的碳汇作用[J]. 科学通报, 2022, 67(15): 1715−1726. doi: 10.1360/TB-2021-0672Xie Shucheng, Jiao Nianzhi, Luo Genming, et al. Evolution of biotic carbon pumps in Earth history: microbial roles as a carbon sink in oceans[J]. Chinese Science Bulletin, 2022, 67(15): 1715−1726. doi: 10.1360/TB-2021-0672 [2] 焦念志. 从戈登科学前沿论坛看海洋碳汇研究动向[C]. 上海: 第四届地球系统科学大会摘要, 2016.Jiao Nianzhi. On the the tendency of the tendency of ocean carbon sequestration from Gordon research conferences[C]. Shanghai: Abstract of the 4th Confeference on Earth System Science, 2016. [3] 孙军. 海洋碳中和途径知多少?[J]地球科学, 2022, 47(10): 3927−3928.Sun Jun. How many pathways we have for the marine carbon neutrality?[J]. Earth Science, 2022, 47(10): 3927−3928. [4] 孙军, 李晓倩, 陈建芳, 等. 海洋生物泵研究进展[J]. 海洋学报, 2016, 38(4): 1−21. doi: 10.3969/j.issn.0253-4193.2016.04.001Sun Jun, Li Xiaoqian, Chen Jianfang, et al. Progress in oceanic biological pump[J]. Haiyang Xuebao, 2016, 38(4): 1−21. doi: 10.3969/j.issn.0253-4193.2016.04.001 [5] Jiao Nianzhi, Herndl G J, Hansell D A, et al. Microbial production of recalcitrant dissolved organic matter: long-term carbon storage in the global ocean[J]. Nature Reviews Microbiology, 2010, 8(8): 593−599. doi: 10.1038/nrmicro2386 [6] Jiao Nianzhi, Cai Ruanhong, Zheng Qiang, et al. Unveiling the enigma of refractory carbon in the ocean[J]. National Science Review, 2018, 5(4): 459−463. doi: 10.1093/nsr/nwy020 [7] 焦念志, 戴民汉, 翦知湣, 等. 海洋储碳机制及相关生物地球化学过程研究策略[J]. 科学通报, 2022, 67(15): 1600−1606. doi: 10.1360/TB-2022-0057Jiao Nianzhi, Dai Minhan, Jian Zhimin, et al. Research strategies for ocean carbon storage mechanisms and effects[J]. Chinese Science Bulletin, 2022, 67(15): 1600−1606. doi: 10.1360/TB-2022-0057 [8] 张兴亮. 海洋惰性溶解有机碳库与海侵黑色页岩[J]. 科学通报, 2022, 67(15): 1607−1613. doi: 10.1360/TB-2021-0613Zhang Xingliang. Marine refractory dissolved organic carbon and transgressive black shales[J]. Chinese Science Bulletin, 2022, 67(15): 1607−1613. doi: 10.1360/TB-2021-0613 [9] Jiao Nianzhi, Wang Hong, Xu Guanhua, et al. Blue carbon on the rise: challenges and opportunities[J]. Naional Science Review, 2018, 5(4): 464−468. [10] 李三忠, 刘丽军, 索艳慧, 等. 碳构造: 一个地球系统科学新范式[J]. 科学通报, 2023, 68(4): 309−338. doi: 10.1360/TB-2022-0741Li Sanzhong, Liu Lijun, Suo Yanhui, et al. Carbon tectonics: a new paradigm for Earth system science[J]. Chinese Science Bulletin, 2023, 68(4): 309−338. doi: 10.1360/TB-2022-0741 [11] Volkman J K, Barrett S M, Blackburn S I, et al. Microalgal biomarkers: a review of recent research developments[J]. Organic Geochemistry, 1998, 29(5/7): 1163−1179. [12] Higginson M J, Altabet M A. Initial test of the silicic acid leakage hypothesis using sedimentary biomarkers[J]. Geophysical Research Letters, 2004, 31(18): L18303. [13] Xie Shucheng, Pancost R D, Yin Hongfu, et al. Two episodes of microbial change coupled with Permo/Triassic faunal mass extinction[J]. Nature, 2005, 434(7032): 494−497. doi: 10.1038/nature03396 [14] 白有成, 陈建芳, 李宏亮, 等. 楚科奇海附近表层沉积物中类脂生物标志物的分布特征和意义[J]. 海洋学报, 2010, 32(2): 106−117.Bai Youcheng, Chen Jianfang, Li Hongliang, et al. The distribution of lipids biomarkers in the surface sediments of the Chukchi Sea and their implications[J]. Haiyang Xuebao, 2010, 32(2): 106−117. [15] 郭新宇, 张海龙, 李莉, 等. 近30年来东海长江口泥质区浮游植物生产力与群落结构变化的生物标志物记录[J]. 中国海洋大学学报, 2020, 50(2): 85−94Guo Xinyu, Zhang Hailong, Li Li, et al. Biomarker records of phytoplankton productivity and community structure changes in the mud area of the Yangtze River Estuary in the East China Sea during the last 30 years[J]. Periodical of Ocean University of China, 2020, 50(2): 85−94. [16] Schouten S, Hopmans E C, Schefub E, et al. Distributional variations in marine crenarchaeotal a new tool for reconstructing ancient sea water temperatures?[J]. Earth and Planetary Science Letters, 2002, 204: 265−274. [17] Wang H Y, He Y K, Liu W G, et al. Lake water depth controlling archaeal tetraether distributions in midlatitude Asia: implications for paleo lake‐level reconstruction[J]. Geophysical Research Letters, 2019, 46(10): 5274−5283. doi: 10.1029/2019GL082157 [18] Damsté J S S, Rijpstra W I C, Hopmans E C, et al. Intact polar and core glycerol dibiphytanyl glycerol tetraether lipids of Group I. 1a and I. 1b Thaumarchaeota in soil[J]. Applied and Environmental Microbiology, 2012, 78(19): 6866−6874. doi: 10.1128/AEM.01681-12 [19] Kou Qiangqiang, Zhu Liping, Wang Junbo, et al. Archaeal tetraether-inferred hydrological variations of Serling Co(Central Tibet) during the late Quaternary[J]. Global and Planetary Change, 2023, 224: 104113. doi: 10.1016/j.gloplacha.2023.104113 [20] Schouten S, Hopmans E C, Schefuß E, et al. Distributional variations in marine crenarchaeotal membrane lipids: a new tool for reconstructing ancient sea water temperatures?[J]. Earth and Planetary Science Letters, 2002, 204(1/2): 265−274. [21] Kim J H, Van Der Meer J, Schouten S, et al. New indices and calibrations derived from the distribution of crenarchaeal isoprenoid tetraether lipids: implications for past sea surface temperature reconstructions[J]. Geochimica et Cosmochimica Acta, 2010, 74(16): 4639−4654. doi: 10.1016/j.gca.2010.05.027 [22] 陈文深, 于培松, 韩喜彬, 等. 南极罗斯海表层沉积物GDGT含量分布及其环境意义[J]. 海洋学研究, 2019, 37(1): 30−39. doi: 10.3969/j.issn.1001-909X.2019.01.005Chen Wenshen, Yu Peisong, Han Xibin, et al. Contents and distribution of GDGTs in surface sediments of Ross Sea, Antarctic and their environmental significances[J]. Journal of Marine Sciences, 2019, 37(1): 30−39. doi: 10.3969/j.issn.1001-909X.2019.01.005 [23] Rampen S W, Schouten S, Koning E, et al. A 90kyr upwelling record from the northwestern Indian Ocean using a novel long-chain diol index[J]. Earth and Planetary Science Letters, 2008, 276(1/2): 207−213. [24] Rampen S W, Willmott V, Kim J H, et al. Evaluation of long chain 1, 14-alkyl diols in marine sediments as indicators for upwelling and temperature[J]. Organic Geochemistry, 2014, 76: 39−47. doi: 10.1016/j.orggeochem.2014.07.012 [25] Yang Huan, Xiao Wenjie, Słowakiewicz M, et al. Depth-dependent variation of archaeal ether lipids along soil and peat profiles from southern China: implications for the use of isoprenoidal GDGTs as environmental tracers[J]. Organic Geochemistry, 2019, 128: 42−56. doi: 10.1016/j.orggeochem.2018.12.009 [26] Kuypers M M M, Blokker P, Erbacher J, et al. Massive expansion of marine archaea during a Mid-Cretaceous oceanic anoxic event[J]. Science, 2001, 293(5527): 92−95. doi: 10.1126/science.1058424 [27] Schouten S, Hopmans E C, Forster A, et al. Extremely high sea-surface temperatures at low latitudes during the middle Cretaceous as revealed by archaeal membrane lipids[J]. Geology, 2003, 31(12): 1069−1072. doi: 10.1130/G19876.1 [28] Jenkyns H C, Schouten-Huibers L, Schouten S, et al. Warm Middle Jurassic-Early Cretaceous high-latitude sea-surface temperatures from the Southern Ocean[J]. Climate of the Past, 2012, 8(1): 215−226. doi: 10.5194/cp-8-215-2012 [29] Roshan S, Devries T. Efficient dissolved organic carbon production and export in the oligotrophic ocean[J]. Nature Communications, 2017, 8(1): 2036. doi: 10.1038/s41467-017-02227-3 [30] 陈立奇. 极区海洋碳池变化性和脆弱性及其探测工程技术[J]. 中国工程科学, 2009, 11(10): 79−85. doi: 10.3969/j.issn.1009-1742.2009.10.012Chen Liqi. Variability and vulnerability of carbon pools in Southern Ocean and western Arctic Ocean and their exploratory engineering technology[J]. Strategic Study of Chinese Academy of Engineerng, 2009, 11(10): 79−85. doi: 10.3969/j.issn.1009-1742.2009.10.012 [31] Le Quéré C, Orr J C, Monfray P, et al. Interannual variability of the oceanic sink of CO2 from 1979 through 1997[J]. Global Biogeochemical Cycles, 2000, 14(4): 1247−1265. doi: 10.1029/1999GB900049 [32] 张凡, 高众勇, 孙恒. 南极普里兹湾碳循环研究进展[J]. 极地研究, 2013, 25(3): 284−293.Zhang Fan, Gao Zhongyong, Sun Heng. Advances in carbon-cycle research for Prydz Bay, the Antarctica[J]. Chinese Journal of Polar Research, 2013, 25(3): 284−293. [33] Yang Dan, Han Zhengbing, Han Xibin, et al. Biomarker records of D5-6 columns in the eastern Antarctic Peninsula waters: responses of planktonic communities and bio-pump structures to sea ice global warming in the past centenary[J]. Advances in Polar Science, 2021, 32(1): 28−41. [34] Stammerjohn S E, Martinson D G, Smith R C, et al. Trends in Antarctic annual sea ice retreat and advance and their relation to El Niño–Southern Oscillation and Southern Annular Mode variability[J]. Journal of Geophysical Research: Oceans, 2008, 113(C3): C03S90. [35] Stammerjohn S, Massom R, Rind D, et al. Regions of rapid sea ice change: an inter-hemispheric seasonal comparison[J]. Geophysical Research Letters, 2012, 39(6): L06501. [36] Orr J C, Fabry V J, Aumont O, et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms[J]. Nature, 2005, 437(7059): 681−686. doi: 10.1038/nature04095 [37] Matson P G, Martz T R, Hofmann G E. High-frequency observations of pH under Antarctic sea ice in the southern Ross Sea[J]. Antarctic Science, 2011, 23(6): 607−613. doi: 10.1017/S0954102011000551 [38] Smith Jr W O, Shields A R, Dreyer J C, et al. Interannual variability in vertical export in the Ross Sea: magnitude, composition, and environmental correlates[J]. Deep Sea Research Part I: Oceanographic Research Papers, 2011, 58(2): 147−159. doi: 10.1016/j.dsr.2010.11.007 [39] Pusceddu A, Cattaneo-Vietti R, Albertelli G, et al. Origin, biochemical composition and vertical flux of particulate organic matter under the pack ice in Terra Nova Bay(Ross Sea, Antarctica) during late summer 1995[J]. Polar Biology, 1999, 22(2): 124−132. doi: 10.1007/s003000050399 [40] Collier R, Dymond J, Honjo S, et al. The vertical flux of biogenic and lithogenic material in the Ross Sea: moored sediment trap observations 1996-1998[J]. Deep Sea Research Part II: Topical Studies in Oceanography, 2000, 47(15/16): 3491−3520. [41] Langone L, Frignani M, Ravaioli M, et al. Particle fluxes and biogeochemical processes in an area influenced by seasonal retreat of the ice margin(northwestern Ross Sea, Antarctica)[J]. Journal of Marine Systems, 2000, 27(1/3): 221−234. [42] Murphy E J, Watkins J L, Trathan P N, et al. Spatial and temporal operation of the Scotia Sea ecosystem: a review of large-scale links in a krill centred food web[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2007, 362(1477): 113−148. doi: 10.1098/rstb.2006.1957 [43] Mari X, Passow U, Migon C, et al. Transparent exopolymer particles: effects on carbon cycling in the ocean[J]. Progress in Oceanography, 2017, 151: 13−37. doi: 10.1016/j.pocean.2016.11.002 [44] Wu Shuangye, Hou Shugui. Impact of icebergs on net primary productivity in the Southern Ocean[J]. The Cryosphere, 2017, 11(2): 707−722. doi: 10.5194/tc-11-707-2017 [45] Ducklow H W, Wilson S E, Post A F, et al. Particle flux on the continental shelf in the Amundsen Sea Polynya and Western Antarctic Peninsula[J]. Elementa: Science of the Anthropocene, 2015, 3(1): 000046. [46] Smith Jr W O, Sedwick P N, Arrigo K R, et al. The Ross Sea in a sea of change[J]. Oceanography, 2012, 25(3): 90−103. doi: 10.5670/oceanog.2012.80 [47] 陈建芳. 南海沉降颗粒物的生物地球化学过程及其在古环境研究中的意义[D]. 上海: 同济大学, 2005.Chen Jianfang. Biogeochemistry of settling particles in the South China Sea and its significance for paleo-environment studies[D]. Shanghai: Tongji University, 2005. [48] Cañadas F, Papineau D, Algeo T J, et al. Upwelling-driven high organic production in the late Ediacaran[J]. Communications Earth & Environment, 2024, 5(1): 461. [49] 周郑鹏, 肖文申, 王汝建, 等. 南极罗斯海表层沉积物中生源组分的分布规律及其环境指示意义[J]. 海洋地质与第四纪地质, 2022, 42(4): 12−23.Zhou Zhengpeng, Xiao Wenshen, Wang Rujian, et al. Distribution patterns of biogenic components in surface sediments of the Ross Sea and their environmental implications[J]. Marine Geology & Quaternary Geology, 2022, 42(4): 12−23. [50] 宋乐慧, 韩喜彬, 李家彪, 等. 罗斯海西部末次冰盛期以来沉积环境重建: 有机碳与生物标志化合物分析[J]. 海洋学报, 2019, 41(9): 52−64. doi: 10.3969/j.issn.0253-4193.2019.09.005Song Lehui, Han Xibin, Li Jiabiao, et al. Western Ross Sea sedimentary environment reconstruction since the Last Glacial Maximum based on organic carbon and biomarker analyses[J]. Haiyang Xuebao, 2019, 41(9): 52−64. doi: 10.3969/j.issn.0253-4193.2019.09.005 [51] 韩喜彬, 赵军, 初凤友, 等. 南极半岛东北海域表层沉积有机质来源及其沉积环境[J]. 海洋学报, 2015, 37(8): 26−38. doi: 10.3969/j.issn.0253-4193.2015.08.003Han Xibin, Zhao Jun, Chu Fengyou, et al. The source of organic matter and its sedimentary environment of the bottom surface sediment in Northeast waters to Antarctic Peninsula based on the biomarker features[J]. Haiyang Xuebao, 2015, 37(8): 26−38. doi: 10.3969/j.issn.0253-4193.2015.08.003 [52] Sampei Y, Matsumoto E. C/N ratios in a sediment core from Nakaumi Lagoon, southwest Japan—usefulness as an organic source indicator[J]. Geochemical Journal, 2001, 35(3): 189−205. doi: 10.2343/geochemj.35.189 [53] 黄邦钦, 柳欣. 边缘海浮游生态系统对生物泵的调控作用[J]. 地球科学进展, 2015, 30(3): 385−395. doi: 10.11867/j.issn.1001-8166.2015.03.0385Huang Bangqin, Liu Xin. Review on planktonic ecosystem and its control on biological pump in the marginal seas[J]. Advances in Earth Science, 2015, 30(3): 385−395. doi: 10.11867/j.issn.1001-8166.2015.03.0385 [54] 康建成, 唐述林, 刘雷保. 南极海冰与气候[J]. 地球科学进展, 2005, 20(7): 786−793. doi: 10.3321/j.issn:1001-8166.2005.07.013Kang Jiancheng, Tang Shulin, Liu Leibao. Antarctic sea ice and climate[J]. Advances in Earth Science, 2005, 20(7): 786−793. doi: 10.3321/j.issn:1001-8166.2005.07.013 [55] 马丽娟, 陆龙骅, 卞林根. 南极海冰的时空变化特征[J]. 极地研究, 2004, 16(1): 29−37.Ma Lijuan, Lu Longhua, Bian Lingen. Spatio-temporal character of Antarctic sea ice variation[J]. Chinese Journal of Polar Research, 2004, 16(1): 29−37. [56] Arrigo K R, Van Dijken G L. Annual changes in sea-ice, chlorophyll a, and primary production in the Ross Sea, Antarctica[J]. Deep Sea Research Part II: Topical Studies in Oceanography, 2004, 51(1/3): 117−138. [57] Ficken K J, Li Baocai, Swain D L. An n-alkane proxy for the sedimentary input of submerged/floating freshwater aquatic macrophytes[J]. Organic Geochemistry, 2000, 31(7/8): 745−749. [58] Smith Jr W O, Ainley D G, Arrigo K R, et al. The oceanography and ecology of the Ross Sea[J]. Annual Review of Marine Science, 2014, 6: 469−487. doi: 10.1146/annurev-marine-010213-135114 [59] Cranwell P A. Chain-length distribution of n-alkanes from lake sediments in relation to post-glacial environmental change[J]. Freshwater Biology, 1973, 3(3): 259−265. doi: 10.1111/j.1365-2427.1973.tb00921.x [60] Rielley G, Collier R J, Jones D M, et al. The biogeochemistry of Ellesmere Lake, U. K. —I: source correlation of leaf wax inputs to the sedimentary lipid record[J]. Organic Geochemistry, 1991, 17(6): 901−912. doi: 10.1016/0146-6380(91)90031-E [61] Meyers P A. Applications of organic geochemistry to paleolimnological reconstructions: a summary of examples from the Laurentian Great Lakes[J]. Organic Geochemistry, 2003, 34(2): 261−289. doi: 10.1016/S0146-6380(02)00168-7 [62] Meyers P A, Ishiwatari R. Lacustrine organic geochemistry—an overview of indicators of organic matter sources and diagenesis in lake sediments[J]. Organic Geochemistry, 1993, 20(7): 867−900. doi: 10.1016/0146-6380(93)90100-P [63] 王启栋, 宋金明, 李学刚. 黄河口湿地有机碳来源及其对碳埋藏提升策略的启示[J]. 生态学报, 2015, 35(2): 568−576.Wang Qidong, Song Jinming, Li Xuegang. Sources of organic carbon in the wetlands of the Yellow River estuary and instructions on carbon burial promotion strategies[J]. Acta Ecologica Sinica, 2015, 35(2): 568−576. [64] Meyers P A. Organic geochemical proxies of paleoceanographic, paleolimnologic, and paleoclimatic processes[J]. Organic Geochemistry, 1997, 27(5/6): 213−250. [65] Rosenthal Y, Dahan M, Shemesh A. Southern ocean contributions to glacial-interglacial changes of atmospheric pCO2: an assessment of carbon isotope records in diatoms[J]. Paleoceanography, 2000, 15(1): 65−75. doi: 10.1029/1999PA000369 [66] Philip R P, Gilbert T D. Characterization of petroleum source rocks and shales by pyrolysis-gas chromatography-mass spectrometry-multiple ion detection[J]. Organic Geochemistry, 1984, 6: 489−501. doi: 10.1016/0146-6380(84)90072-X [67] Nichols P D, Palmisano A C, Volkman J K, et al. Occurrence of an isoprenoid C25 diunasaturated alkene and high neutral lipid content in antractic sea-ice diatom communities[J]. Journal of Phycology, 1988, 24(1): 90−96. doi: 10.1111/j.1529-8817.1988.tb04459.x [68] 吴庆余, 殷实, 盛国英, 等. 发现于浮游硅藻中的长链正烷烃[J]. 科学通报, 1992, 37(24): 2266−2269.Wu Qingyu, Yin Shi, Sheng Guoying, et al. The long chain n-alkanes are found in floating diatoms[J]. Chinese Science Bulletin, 1992, 37(24): 2266−2269. [69] Smith Jr W O, Dinniman M S, Hofmann E E, et al. The effects of changing winds and temperatures on the oceanography of the Ross Sea in the 21st century[J]. Geophysical Research Letters, 2014, 41(5): 1624−1631. doi: 10.1002/2014GL059311 [70] DeMaster D J, Ragueneau O, Nittrouer C A. Preservation efficiencies and accumulation rates for biogenic silica and organic C, N, and P in high-latitude sediments: The Ross Sea[J]. Journal of Geophysical Research: Oceans, 1996, 101(C8): 18501−18518. doi: 10.1029/96JC01634 [71] Goffart A, Catalano G, Hecq J H. Factors controlling the distribution of diatoms and Phaeocystis in the Ross Sea[J]. Journal of Marine Systems, 2000, 27(1/3): 161−175. [72] Zimmerman A R, Cornelissen G. Consider fjord-assisted carbon storage[J]. Environmental Science & Technology, 2018, 52(19): 10911−10913. [73] Dewar W K, Bingham R J, Iverson R L, et al. Does the marine biosphere mix the ocean?[J]. Journal of Marine Research, 2006, 64(4): 541−561. doi: 10.1357/002224006778715720 [74] Volkman J K, Maxwell J R. Acyclic isoprenoids as biological markers[M]//Johns R B. Biological Markers in the Sedimentary Record. New York: Elsevier, 1986: 1−42. [75] Didyk B M, Simoneit B R T, Brassell S C, et al. Organic geochemical indicators of palaeoenvironmental conditions of sedimentation[J]. Nature, 1978, 272(5650): 216−222. doi: 10.1038/272216a0 [76] 傅家谟, 盛国英, 许家友, 等. 应用生物标志化合物参数判识古沉积环境[J]. 地球化学, 1991, 20(1): 1−12. doi: 10.3321/j.issn:0379-1726.1991.01.001Fu Jiamo, Sheng Guoying, Xu Jiayou, et al. Application of biomarker compounds in assessment of paleoenvironments of Chinese terrestrial sediments[J]. Geochimica, 1991, 20(1): 1−12. doi: 10.3321/j.issn:0379-1726.1991.01.001 [77] Mangelsdorf K, Rullkötter J. Natural supply of oil-derived hydrocarbons into marine sediments along the California continental margin during the late Quaternary[J]. Organic Geochemistry, 2003, 34(8): 1145−1159. doi: 10.1016/S0146-6380(03)00059-7 [78] 董宏坤, 万世明, 刘喜停. 海洋沉积物早期成岩作用研究进展[J]. 沉积学报, 2022, 40(5): 1172−1187.Dong Hongkun, Wan Shiming, Liu Xiting. Research progress on geochemical behavior of minerals and elements in early diagenesis of marine sediments[J]. Acta Sedimentologica Sinica, 2022, 40(5): 1172−1187. [79] Frignani M, Giglio F, Accornero A, et al. Sediment characteristics at selected sites of the Ross Sea continental shelf: does the sedimentary record reflect water column fluxes?[J]. Antarctic Science, 2003, 15(1): 133−139. doi: 10.1017/S0954102003001123 [80] Belt S T, Smik L, Brown T A, et al. Source identification and distribution reveals the potential of the geochemical Antarctic sea ice proxy IPSO25[J]. Nature Communications, 2016, 7(1): 12655. doi: 10.1038/ncomms12655 [81] Wakeham S G, Hopmans E C, Schouten S, et al. Archaeal lipids and anaerobic oxidation of methane in euxinic water columns: a comparative study of the Black Sea and Cariaco Basin[J]. Chemical Geology, 2004, 205(3/4): 427−442. [82] Schouten S, Hopmans E C, Baas M, et al. Intact membrane lipids of “Candidatus Nitrosopumilus maritimus, ” a cultivated representative of the cosmopolitan mesophilic group I crenarchaeota[J]. Applied and Environmental Microbiology, 2008, 74(8): 2433−2440. [83] Shen Yuan, Guilderson T P, Chavez F P, et al. Important contribution of bacterial carbon and nitrogen to sinking particle export[J]. Geophysical Research Letters, 2023, 50(11): e2022GL102485. doi: 10.1029/2022GL102485 [84] Zheng Yanhong, Singarayer J S, Cheng Peng, et al. Holocene variations in peatland methane cycling associated with the Asian summer monsoon system[J]. Nature Communications, 2014, 5(1): 4631. doi: 10.1038/ncomms5631 [85] Hu Jianfang, Zhou Haoda, Peng Ping'an, et al. Seasonal variability in concentrations and fluxes of glycerol dialkyl glycerol tetraethers in Huguangyan Maar Lake, SE China: Implications for the applicability of the MBT-CBT paleotemperature proxy in lacustrine settings[J]. Chemical Geology, 2016, 420: 200−212. doi: 10.1016/j.chemgeo.2015.11.008 [86] Woese C R, Kandler O, Wheelis M L. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya[J]. Proceedings of the National Academy of Sciences of the United Stated of America, 1990, 87(12): 4576−4579. [87] Koga Y, Nishihara M, Morii H, et al. Ether polar lipids of methanogenic bacteria: structures, comparative aspects, and biosyntheses[J]. Microbiology and Molecular Biology Reviews, 1993, 57(1): 164−182. [88] Schouten S, Forster A, Panoto F E, et al. Towards calibration of the TEX86 palaeothermometer for tropical sea surface temperatures in ancient greenhouse worlds[J]. Organic Geochemistry, 2007, 38(9): 1537−1546. doi: 10.1016/j.orggeochem.2007.05.014 [89] Blaga C I, Reichart G J, Heiri O, et al. Tetraether membrane lipid distributions in water-column particulate matter and sediments: a study of 47 European lakes along a north-south transect[J]. Journal of Paleolimnology, 2009, 41(3): 523−540. doi: 10.1007/s10933-008-9242-2 [90] Xiao Wenjie, Xu Yunping, Lin Jian, et al. Global scale production of brGDGTs by benthic marine bacteria: Implication for developing ocean bottom environmental proxies[J]. Global and Planetary Change, 2022, 211: 103783. doi: 10.1016/j.gloplacha.2022.103783 [91] Schouten S, Hopmans E C, Damsté J S S. The organic geochemistry of glycerol dialkyl glycerol tetraether lipids: a review[J]. Organic Geochemistry, 2013, 54: 19−61. doi: 10.1016/j.orggeochem.2012.09.006 [92] Lechtenfeld O J, Hertkorn N, Shen Yuan, et al. Marine sequestration of carbon in bacterial metabolites[J]. Nature Communications, 2015, 6(1): 6711. doi: 10.1038/ncomms7711 [93] Schimel J. Playing scales in the methane cycle: from microbial ecology to the globe[J]. Proceedings of the National Academy of Sciences of United States of America, 2004, 101(34): 12400−12401. doi: 10.1073/pnas.0405075101 [94] 周文广, 阮榕生. 微藻生物固碳技术进展和发展趋势[J]. 中国科学: 化学, 2014, 44(1): 63−78. doi: 10.1360/032013-256Zhou Wenguang, Ruan Rongsheng. Biological mitigation of carbon dioxide via microalgae: recent development and future direction[J]. Scientia Sinica: Chimica, 2014, 44(1): 63−78. doi: 10.1360/032013-256 [95] Kim J H, Schouten S, Hopmans E C, et al. Global sediment core-top calibration of the TEX86 paleothermometer in the ocean[J]. Geochimica et Cosmochimica Acta, 2008, 72(4): 1154−1173. doi: 10.1016/j.gca.2007.12.010 [96] Shevenell A E, Ingalls A E, Domack E W, et al. Holocene Southern Ocean surface temperature variability west of the Antarctic Peninsula[J]. Nature, 2011, 470(7333): 250−254. doi: 10.1038/nature09751 [97] Weijers J W H, Schouten S, Spaargaren O C, et al. Occurrence and distribution of tetraether membrane lipids in soils: Implications for the use of the TEX86 proxy and the BIT index[J]. Organic Geochemistry, 2006, 37(12): 1680−1693. doi: 10.1016/j.orggeochem.2006.07.018 [98] Etourneau J, Collins L G, Willmott V, et al. Holocene climate variations in the western Antarctic Peninsula: evidence for sea ice extent predominantly controlled by changes in insolation and ENSO variability[J]. Climate of the Past, 2013, 9(4): 1431−1446. doi: 10.5194/cp-9-1431-2013 [99] Arrigo K R, Van Dijken G, Long M. Coastal Southern Ocean: a strong anthropogenic CO2 sink[J]. Geophysical Research Letters, 2008, 35(21): L21602. [100] Smith Jr W O, Asper V L. The influence of phytoplankton assemblage composition on biogeochemical characteristics and cycles in the southern Ross Sea, Antarctica[J]. Deep Sea Research Part I: Oceanographic Research Papers, 2001, 48(1): 137−161. doi: 10.1016/S0967-0637(00)00045-5 [101] 于晓果, 边叶萍, 阮小燕, 等. 北冰洋沉积物中四醚脂类来源与TEX86指数初步研究[J]. 海洋地质与第四纪地质, 2015, 35(3): 11−22.Yu Xiaoguo, Bian Yeping, Ruan Xiaoyan, et al. Glycerol dialkyl glyceroltetraethers and TEX86 index in surface sediments of the Arctic ocean and the Bering sea[J], Marine Geology & Quaternary Geology, 2015, 35(3): 11−22. [102] Zhao Meixun, Mercer J L, Eglinton G, et al. Comparative molecular biomarker assessment of phytoplankton paleoproductivity for the last 160 kyr off Cap Blanc, NW Africa[J]. Organic Geochemistry, 2006, 37(1): 72−97. doi: 10.1016/j.orggeochem.2005.08.022 [103] Schubert C J, Villanueva J, Calvert S E, et al. Stable phytoplankton community structure in the Arabian Sea over the past 200, 000 years[J]. Nature, 1998, 394(6693): 563−566. doi: 10.1038/29047 [104] Hopmans E C, Weijers J W H, Schefuß E, et al. A novel proxy for terrestrial organic matter in sediments based on branched and isoprenoid tetraether lipids[J]. Earth and Planetary Science Letters, 2004, 224(1/2): 107−116. [105] Weijers J W H, Schouten S, Van Den Donker J C, et al. Environmental controls on bacterial tetraether membrane lipid distribution in soils[J]. Geochimica et Cosmochimica Acta, 2007, 71(3): 703−713. doi: 10.1016/j.gca.2006.10.003 [106] 傅明珠, 王宗灵, 李艳, 等. 胶州湾浮游植物初级生产力粒级结构及固碳能力研究[J]. 海洋科学进展, 2009, 27(3): 357−366. doi: 10.3969/j.issn.1671-6647.2009.03.009Fu Mingzhu, Wang Zongling, Li Yan, et al. Study on size-fraction and carbon fixation of phytoplankton primary productivity in Jiaozhou bay[J]. Advances in Marine Science, 2009, 27(3): 357−366. doi: 10.3969/j.issn.1671-6647.2009.03.009 [107] Seki O, Ikehara M, Kawamura K, et al. Reconstruction of paleoproductivity in the Sea of Okhotsk over the last 30 kyr[J]. Paleoceanography, 2004, 19(1): PA1016. [108] Siegenthaler U, Sarmiento J L. Atmospheric carbon dioxide and the ocean[J]. Nature, 1993, 365(6442): 119−125. doi: 10.1038/365119a0 [109] Anderson R, Archer D, Bathmann U, et al. A new vision of ocean biogeochemistry after a decade of the Joint Global Ocean Flux Study(JGOFS)[R]. Berlin: Ambio(Special Report), 2001(10): 4−30. [110] Wilson J D, Barker S, Ridgwell A. Assessment of the spatial variability in particulate organic matter and mineral sinking fluxes in the ocean interior: implications for the ballast hypothesis[J]. Global Biogeochemical Cycles, 2012, 26(4): GB4011. [111] 黄邦钦, 胡俊, 柳欣, 等. 全球气候变化背景下浮游植物群落结构的变动及其对生物泵效率的影响[J]. 厦门大学学报(自然科学版), 2011, 50(2): 402−410.Huang Bangqin, Hu Jun, Liu Xin, et al. Review on phytoplankton community structure and its coupling with Biological carbon pump under global climate changes[J]. Journal of Xiamen University(Natural Science), 2011, 50(2): 402−410. [112] Arrigo K R, Robinson D H, Worthen D L, et al. Phytoplankton community structure and the drawdown of nutrients and CO2 in the Southern Ocean[J]. Science, 1999, 283(5400): 365−367. doi: 10.1126/science.283.5400.365 [113] Volkman J K, Farrington J W, Gagosian R B. Marine and terrigenous lipids in coastal sediments from the Peru upwelling region at 15°S: sterols and triterpene alcohols[J]. Organic Geochemistry, 1987, 11(6): 463−477. doi: 10.1016/0146-6380(87)90003-9 [114] Barrett S M, Volkman J K, Dunstan G A, et al. Sterols of 14 species of marine diatoms(Bacillariophyta)[J]. Journal of Phycology, 1995, 31(3): 360−369. doi: 10.1111/j.0022-3646.1995.00360.x [115] Kawaguchi S, Atkinson A, Bahlburg D, et al. Climate change impacts on Antarctic krill behaviour and population dynamics[J]. Nature Reviews Earth & Environment, 2024, 5(1): 43−58. [116] Cavan E L, Belcher A, Atkinson A, et al. The importance of Antarctic krill in biogeochemical cycles[J]. Nature Communications, 2019, 10(1): 4742. doi: 10.1038/s41467-019-12668-7 [117] 黄梦雪, 王汝建, 肖文申, 等. 罗斯海西北陆架(JOIDES海槽)末次冰期以来冰架消融过程及水动力变化[J]. 海洋地质与第四纪地质, 2016, 36(5): 97−108.Huang Mengxue, Wang Rujian, Xiao Wenshen, et al. Retreat process of Ross Ice Shelf and hydrodynamic changes on northwestern Ross continental shelf since the Last Glacial[J]. Marine Geology & Quaternary Geology, 2016, 36(5): 97−108. [118] Anderson J B, Conway H, Bart P J, et al. Ross Sea paleo-ice sheet drainage and deglacial history during and since the LGM[J]. Quaternary Science Reviews, 2014, 100: 31−54. doi: 10.1016/j.quascirev.2013.08.020 [119] Domack E W, Jacobson E A, Shipp S, et al. Late Pleistocene–holocene retreat of the West Antarctic Ice-Sheet system in the Ross Sea: part 2—sedimentologic and stratigraphic signature[J]. Geological Society of America Bulletin, 1999, 111(10): 1517−1536. doi: 10.1130/0016-7606(1999)111<1517:LPHROT>2.3.CO;2 [120] Bassett S E, Milne G A, Mitrovica J X, et al. Ice sheet and solid Earth influences on far-field sea-level histories[J]. Science, 2005, 309(5736): 925−928. doi: 10.1126/science.1111575 [121] Herndl G J, Reinthaler T, Teira E, et al. Contribution of Archaea to total Prokaryotic production in the deep Atlantic Ocean[J]. Applied and Environmental Microbiology, 2005, 71(5): 2303−2309. doi: 10.1128/AEM.71.5.2303-2309.2005 [122] 杨丹, 付全有, 韩正兵, 等. 南极典型海域浮游生物生产力/群落结构对BP/MCP储碳影响及其年代际变率[J]. 海洋学报, 2024, 46(5): 37−56.Yang Dan, Fu Quanyou, Han Zhengbing, et al. Effects of plankton productivity/community structure on BP/MCP carbon storage and their interdecadal variations in a typical Antarctic waters[J]. Haiyang Xuebao, 2024, 46(5): 37−56. [123] 黄邦钦, 邱勇, 陈纪新. 海洋生物泵研究的若干新进展与展望[J]. 应用海洋学学报, 2019, 38(4): 474−483. doi: 10.3969/J.ISSN.2095-4972.2019.04.003Huang Bangqin, Qiu Yong, Chen Jixin. Progress and prospects on the study of marine biological pump[J]. Journal of Applied Oceanography, 2019, 38(4): 474−483. doi: 10.3969/J.ISSN.2095-4972.2019.04.003 [124] Lampitt R S, NoJi T, Von Bodungen B. What happens to zooplankton faecal pellets? Implications for material flux[J]. Marine Biology, 1990, 104(1): 15−23. doi: 10.1007/BF01313152 [125] 李清毅, 张国民, 王新烨, 等. 微藻在蓝碳中的作用机制及影响因素[J]. 安徽农业科学, 2024, 52(3): 71−76. doi: 10.3969/j.issn.0517-6611.2024.03.017Li Qingyi, Zhang Guomin, Wang Xinye, et al. Mechanism of action and influence factor of microalgae in blue carbon[J]. Journal of Anhui Agricultural Sciences, 2024, 52(3): 71−76. doi: 10.3969/j.issn.0517-6611.2024.03.017 [126] Atkinson A, Schmidt K, Fielding S, et al. Variable food absorption by Antarctic krill: relationships between diet, egestion rate and the composition and sinking rates of their fecal pellets[J]. Deep Sea Research Part II: Topical Studies in Oceanography, 2012, 59−60: 147−158. doi: 10.1016/j.dsr2.2011.06.008 [127] 王建民, 钟巍. 晚冰期新仙女木事件的研究历史及现状[J]. 冰川冻土, 1994, 16(4): 371−379.Wang Jianmin, Zhong Wei. The research history and present situation of Younger Dryas Event in the Last Deglaciadon[J]. Journal of Glaciology and Geocryology, 1994, 16(4): 371−379. [128] 吴时国, 陆钧. 南极普里兹湾1.5万年来气候演变的沉积记录[J]. 海洋学报, 1998, 20(1): 65−73.Wu Shiguo, Lu Jun. Paleoclimatic evolution recorded in sediments of the Prydz Bay, Antarctica in 15 000 years[J]. Haiyang Xuebao, 1998, 20(1): 65−73. [129] Xiao Wenshen, Esper O, Gersonde R. Last Glacial-Holocene climate variability in the Atlantic sector of the Southern Ocean[J]. Quaternary Science Reviews, 2016, 135: 115−137. doi: 10.1016/j.quascirev.2016.01.023 [130] Jouzel J, Masson V, Cattani O, et al. A new 27 ky high resolution East Antarctic climate record[J]. Geophysical Research Letters, 2001, 28(16): 3199−3202. doi: 10.1029/2000GL012243 [131] Lamy F, De Pol-Holz R. Paleoceanography, records: Postglacial South Pacific[M]//Elisas S. The Encyclopedia of Quaternary Science. Amsterdam: Elsevier, 2013: 73−85. [132] Bond G, Showers W, Cheseby M, et al. A pervasive millennial-scale cycle in North Atlantic Holocene and glacial climates[J]. Science, 1997, 278(5341): 1257−1266. doi: 10.1126/science.278.5341.1257 [133] 王宁练, 姚檀栋, Thompson L G, 等. 全新世早期强降温事件的古里雅冰芯记录证据[J]. 科学通报, 2002, 47(11): 818−823.Wang Ninglian, Yao Tandong, Thompson L G, et al. Evidence for Early Holocene Cold event from Gurya ice core records[J]. Chinese Science Bulletin, 2002, 47(11): 818−823. [134] 王绍武. 全新世气候变化[M]. 北京: 气象出版社, 2011.Wang Shaowu. Holocene Climate Change[M]. Beijing: China Meteorological Press, 2011. [135] 施雅风, 孔昭宸, 王苏民, 等. 中国全新世大暖期的气候波动与重要事件[J]. 中国科学 B辑, 1992, 22(12): 1300−1308.Shi Yafeng, Kong Zhaochen, Wang Sumin, et al. Climatic fluctuations and important events during the Holocene Great Warm Period in China[J]. Scientia Sinica, 1992, 22(12): 1300−1308. [136] Moros M, Emeis K, Risebrobakken B, et al. Sea surface temperatures and ice rafting in the Holocene North Atlantic: climate influences on northern Europe and Greenland[J]. Quaternary Science Reviews, 2004, 23(20/22): 2113−2126. [137] Magny M, Haas J N. A major widespread climatic change around 5300 cal. yr BP at the time of the alpine Iceman[J]. Journal Quaternary Science, 2004, 19(5): 423−430. doi: 10.1002/jqs.850 [138] 谢树成, 焦念志, 汪品先. 加强海洋生物碳泵地质演化的研究[J]. 科学通报, 2022, 67(15): 1579−1599.Xie Shucheng, Jiao Nianzhi, Wang Pinxian. Promoting studies on the geological evolution of marine biological carbon pumps[J]. Chinese Science Bulletin, 2022, 67(15): 1579−1599. -




 下载:
下载:


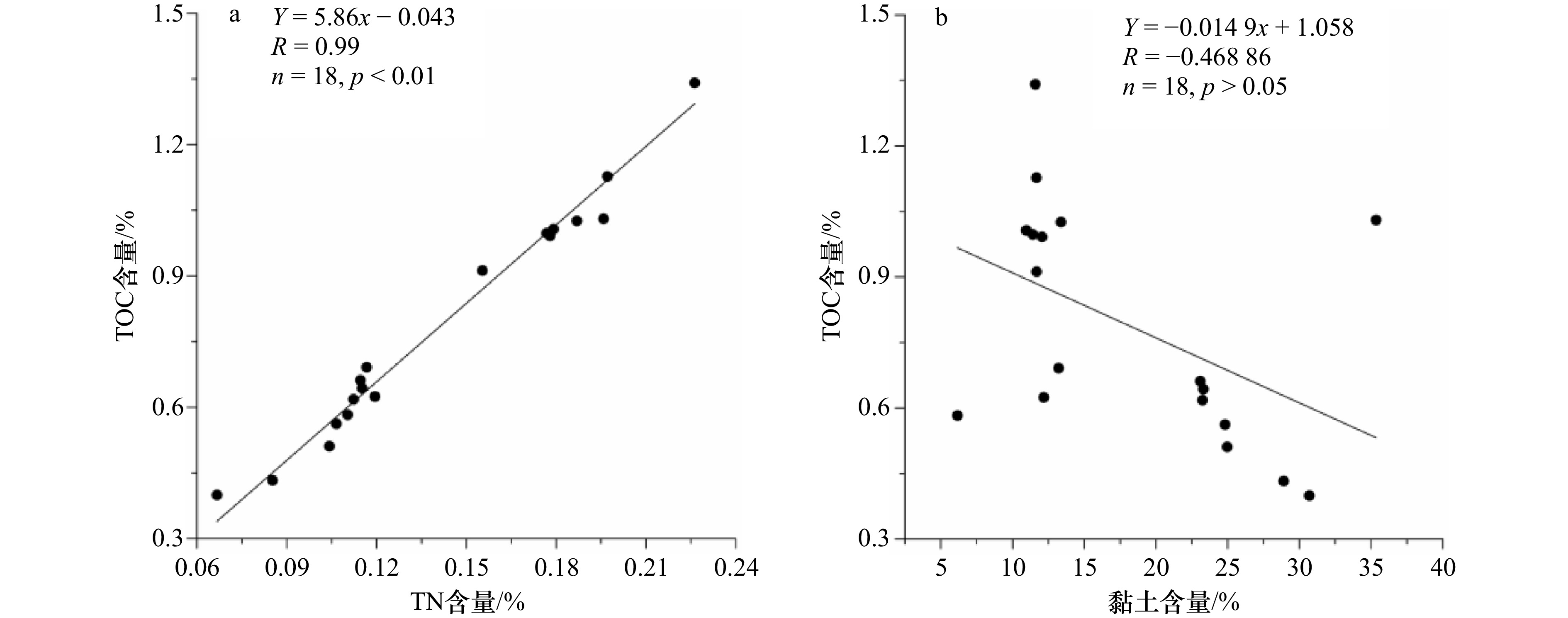




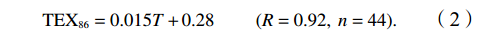
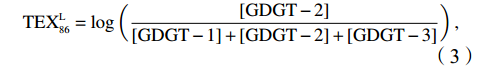



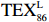



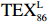


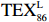





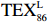
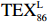
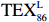
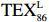
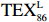
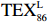
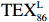
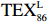

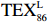

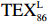



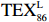









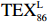

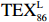
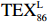

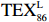



计量
- 文章访问数: 105
- HTML全文浏览量: 51
- PDF下载量: 11
- 被引次数: 0
