

The impacts of submergence duration and salinity on the germination and growth process of Aegiceras corniculatum radicles
-
摘要: 潮位与盐度是影响桐花树幼苗生长发育的重要环境因子,在一定程度上决定桐花树胚胎能否成功着床与快速成苗。本文采用室内控制实验观测桐花树胚胎萌动期根系发育过程,进而探讨盐度、淹水时长、盐度−淹水时长交互作用对胚根萌发及生长的影响。结果表明:(1)模拟低低潮位淹水环境的完全浸泡胚胎24 h/d培养处理组,桐花树胚胎无法萌根。模拟中低潮位淹水环境的完全浸泡胚胎6 h/d + 浸泡胚根18 h/d培养处理组,胚胎平均萌根率最高,胚根最长。模拟高高潮位胚胎基部浸泡水体环境的浸泡胚根培养处理组,胚胎萌根率与平均萌根率最低,胚根数量最少。模拟高高潮位胚胎基部扎入潮滩环境的蛭石浅插培养处理组,胚根萌发最慢,萌根率最高,胚根最短、数量最多。(2)在盐度为0的环境中,培养13 d后胚根最长。在盐度为10的环境中,培养后的前11 d胚根最长、数量最多。在盐度为20的环境中,萌根率、平均萌根率以及胚根长度和数量明显偏低,胚根萌发与生长受到抑制。(3)淹水时长−盐度交互作用对桐花树胚根生长影响显著,若淹水时长、盐度处于桐花树胚胎萌根耐受范围内,淹水时长−盐度复合胁迫下胚胎依然能够正常萌根。人工培育桐花树幼苗,采用完全浸泡胚胎6 h/d + 浸泡胚根18 h/d,盐度为10培养胚根生长发育最优。研究结果可为潮间带桐花树幼苗人工培育与保护提供部分理论依据。Abstract: Tide level and salinity are crucial environmental factors that significantly impact the growth and development of Aegiceras corniculatum seedlings and to a certain extent, determine if their embryos could land successfully and develop into seedling rapidly. In this paper, the process of A. corniculatum embryos developing to seedings was observed by the indoor controlled experiments, during which, the effects of salinity, submergence duration, as well as the interaction between salinity and submergence duration on the germination and growth of radicles were discussed. The results showed that: (1) the embryos of the A. corniculatum were unable to sprout in the treatment group that the embryos were soaked completely for 24 h/d in a simulated lower low tidal level of flooded environment. For the treatment group that the embryos were soaked completely for 6 h/d and the radicles soaked for 18 h/d in a simulated low to medium tidal level of flooded environment, the highest mean rooting rates and the longest radicle of the embryos were produced. In the treatment group that the base of embryos were soaked in a simulated higher high tide flooded environment, the rooting rates and mean rooting rates of embryos were the lowest, accompanied by the least quantities of radicles. However, in the treatment group where the vermiculites were shallowly inserted into the bottom, which simulated a scene that the base of embryo was immersed within the tidal environment, it was found that the radicle germination was the slowest with the highest rooting rates, and the radicles were the shortest, along with the largest quantities. (2) In the salinity environment of 0, the radicles were the longest after a 11 days cultivation. In the salinity environment of 10, highest levels in both the length and quantities of radicles were observed in the first 13 days of embryo culture. In contrast, the rooting rate and the mean rooting rate of the embryos, as well as the length and quantity of radicle were significantly smaller in the salinity environment of 20, which indicated the germination and growth of radicle were inhibited in such salinity environment. (3) The interaction between submergence duration and salinity had a significant impact on the growth of the radicles of A. corniculatum. If both the submergence duration and salinity were within the tolerance ranges of the radicles of A. corniculatum, its embryos could still sprout normally under the stresses of different combination of submergence duration and salinity. Hence, the optimal way to artificially cultivatethe A. corniculatum seedlings, was that the embryo was completely soaked for 6 h/d and the radicle was soaked for 18 h/d in the salinity environment of 10. The study can provide some theoretical basis for the artificial cultivation and protection of A. corniculatum seedlings in the intertidal zone.
-
Key words:
- Aegiceras corniculatum /
- submergence duration /
- salinity /
- radicle
-
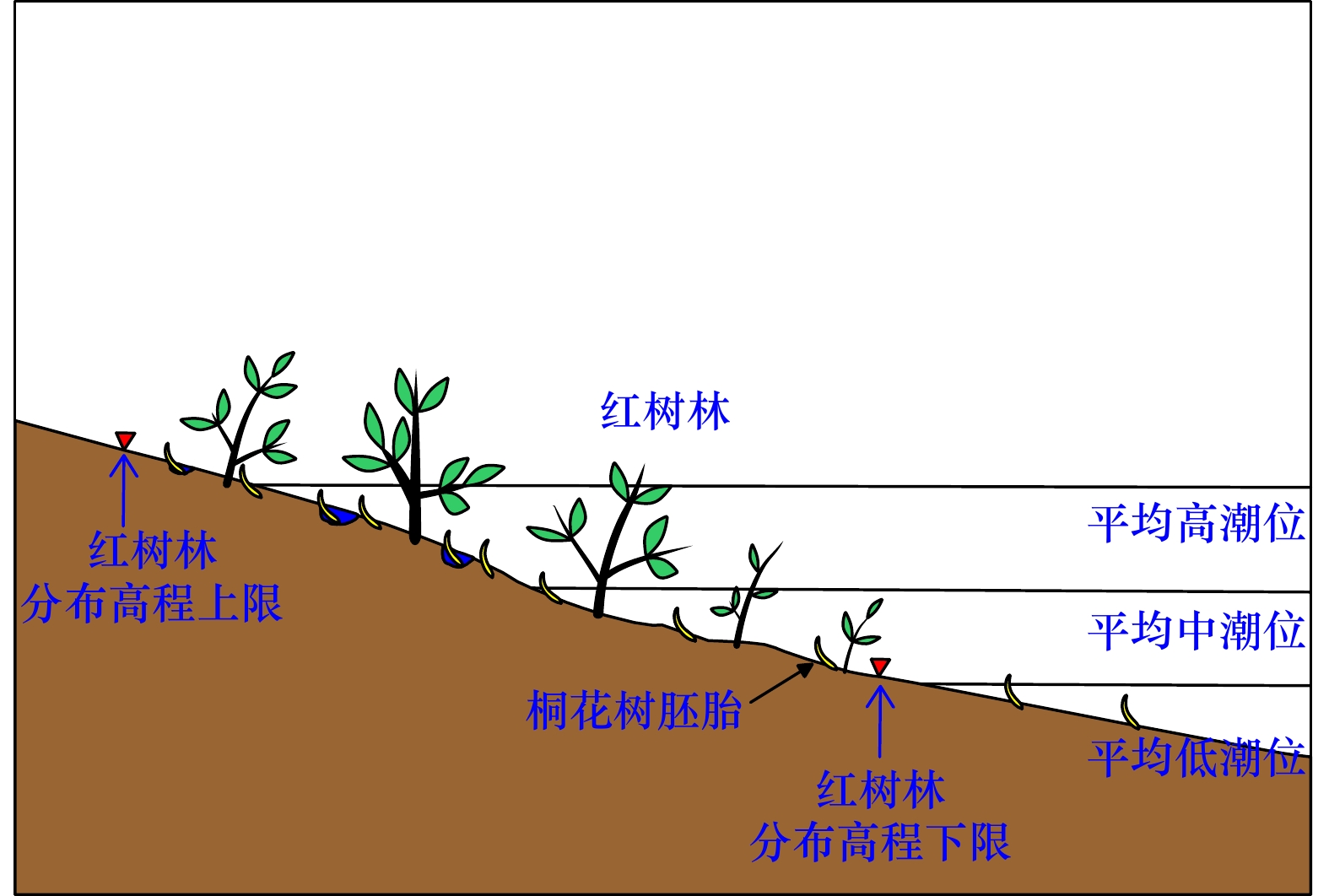
图 1 桐花树胚胎在潮滩上的分布状态
Fig. 1 The distribution of embryos of Aegiceras corniculatum on tidal flat

图 2 室内培养观测
a. 桐花树胚胎室内培养试验设计;b. 胚胎萌根观察
Fig. 2 Observation of indoor culture
a. Design of laboratory culture experiment for the embryos of Aegiceras corniculatum embryo; b. observation of embryo germination
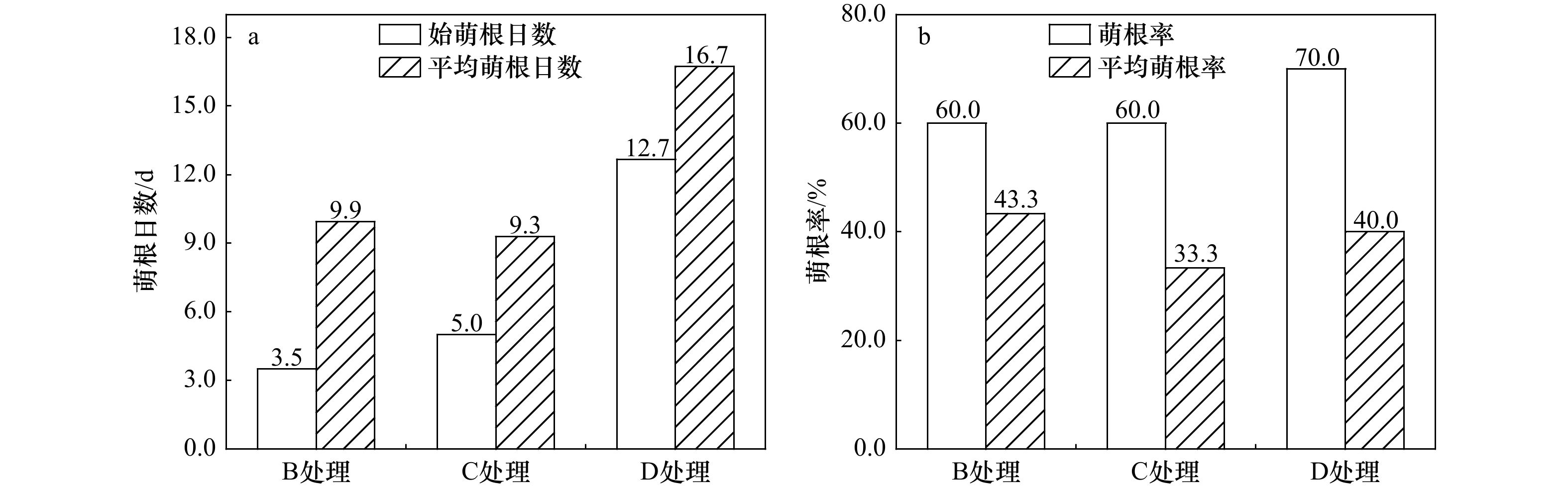
图 3 淹水处理的桐花树胚胎萌根状态
Fig. 3 The state of sprouting roots of Aegiceras corniculatum in flooded treatment
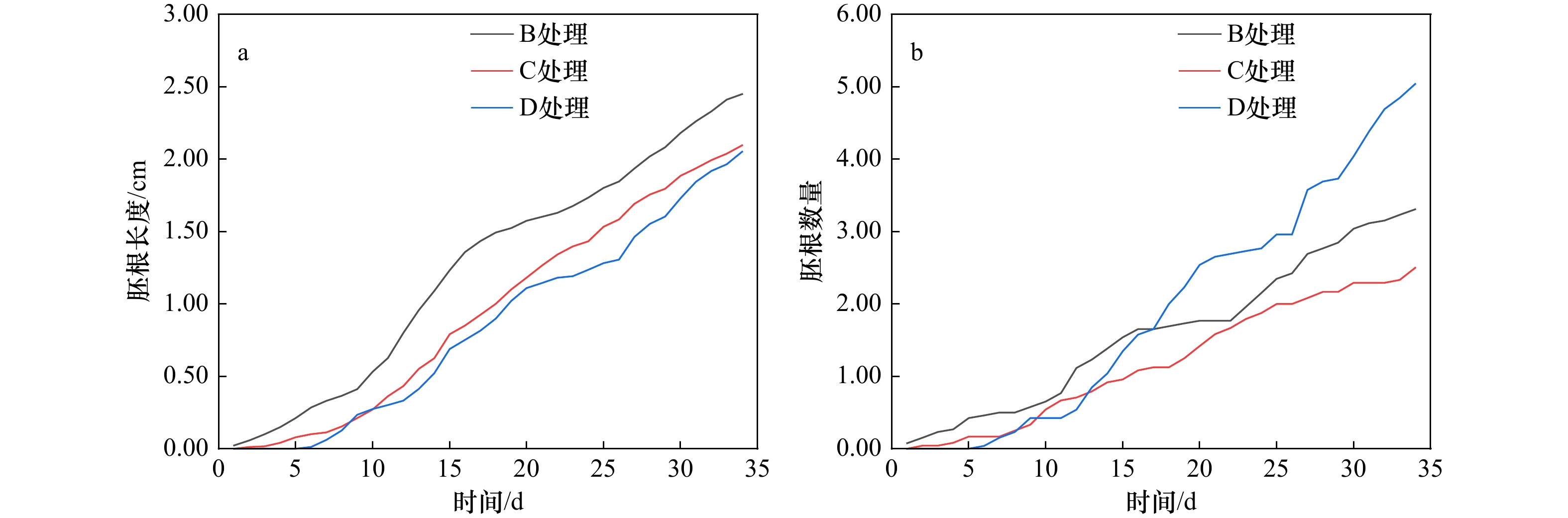
图 4 淹水处理的桐花树胚根生长状态
胚根数量指同一处理胚胎萌根的平均数量,胚根长度指同一处理胚胎全部根系的平均长度
Fig. 4 Growth state of radicle of Aegiceras corniculatum in flooded treatment
The number of radicle refers to the average number of embryonic roots in the same treatment, and the length of radicle refers to the average length of all roots in the same treatment

图 5 盐度处理的桐花树胚胎萌根状态
Fig. 5 The state of sprouting roots of Aegiceras corniculatum in salinity treatment
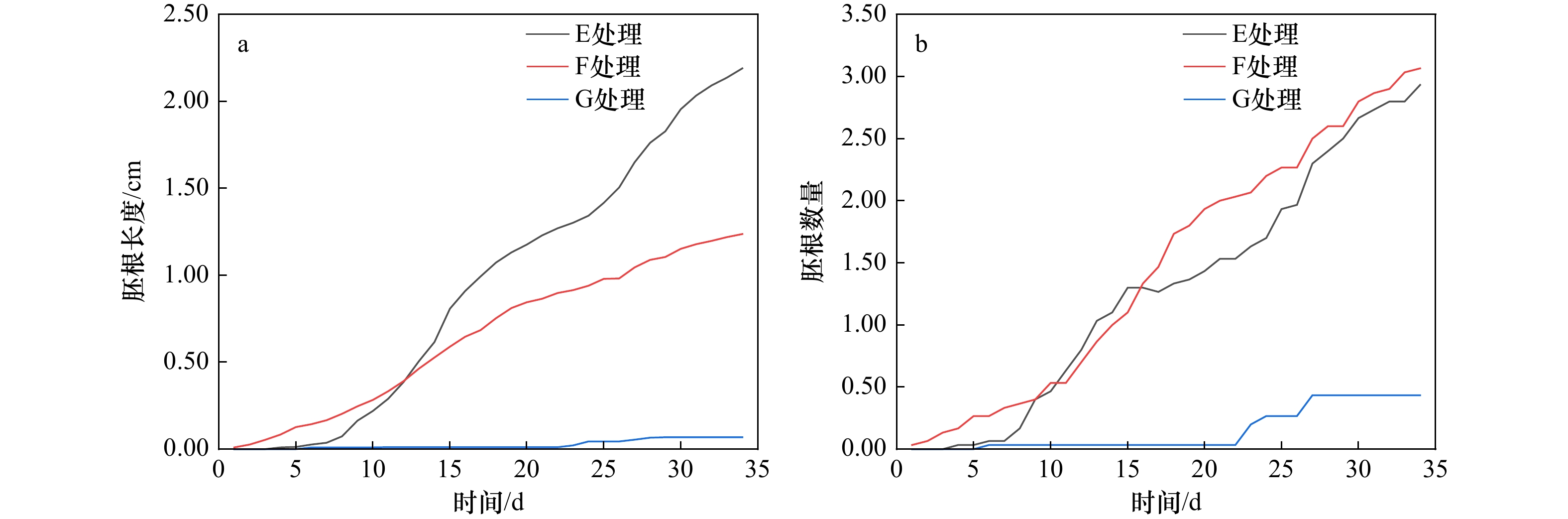
图 6 盐度处理的桐花树胚根生长状态
胚根数量指同一处理胚胎萌根的平均数量,胚根长度指同一处理胚胎全部根系的平均长度
Fig. 6 Growth state of radicle of Aegiceras corniculatum in salinity treatment
The number of radicle refers to the average number of embryonic roots in the same treatment, and the length of radicle refers to the average length of all roots in the same treatment
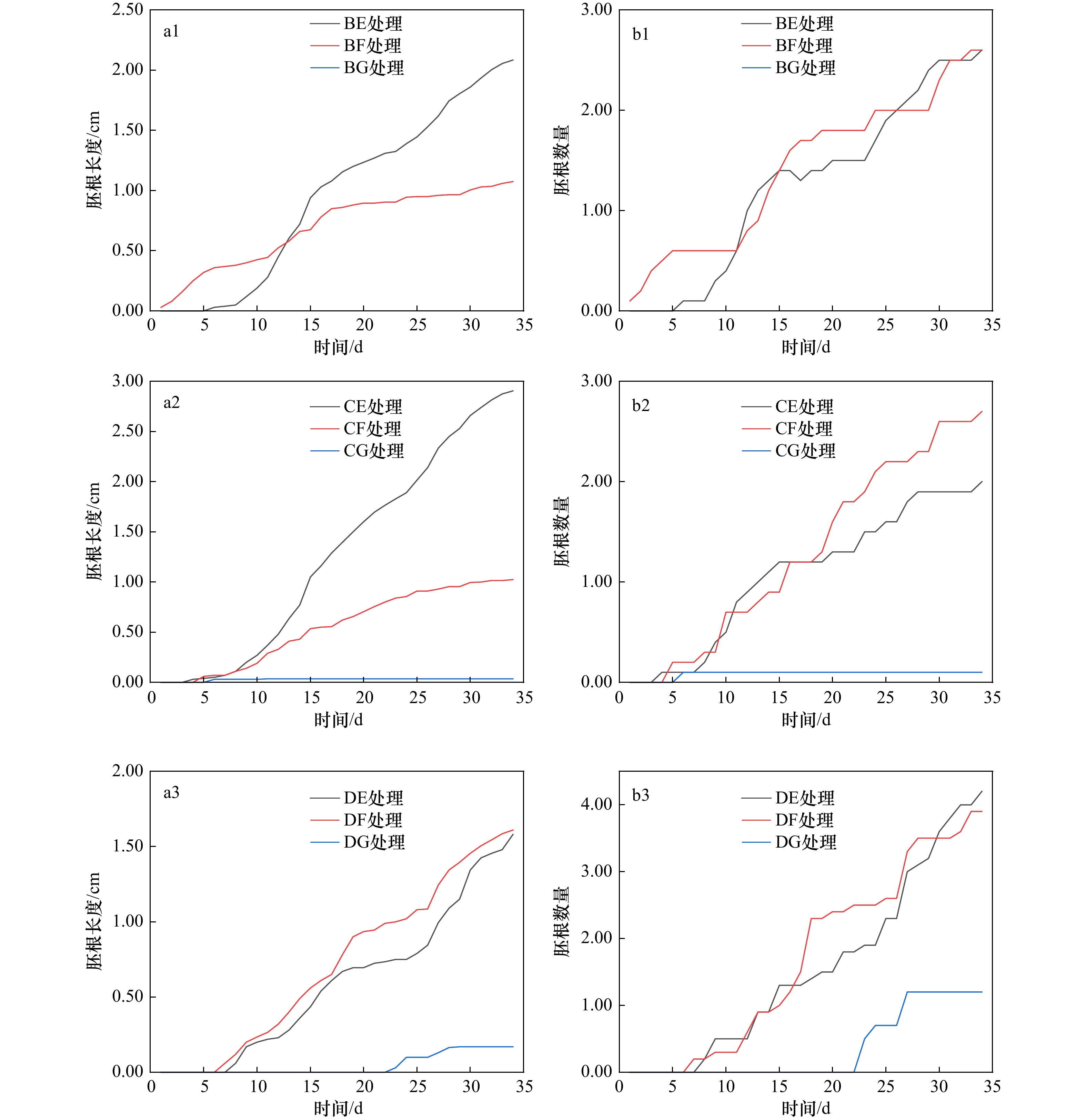
图 7 淹水时长−盐度交互处理下胚根生长过程
胚根数量指同一处理胚胎萌根的平均数量,胚根长度指同一处理胚胎全部根系的平均长度
Fig. 7 Interactive treatment of submergence duration and salinity on the growth process of embryonic roots
The number of radicle refers to the average number of embryonic roots in the same treatment, and the length of radicle refers to the average length of all roots in the same treatment
表 1 室内培养盐度梯度试验设计
Tab. 1 Salinity gradient test design for indoor culture
编号 盐度 处理方式 E 0 自来水静放24 h F 10 盐度为10.9海水加自来水调配 G 20 盐度为10.9海水加海盐调配  下载: 导出CSV
下载: 导出CSV
表 2 淹水处理桐花树胚根长度及数量差异性分析
Tab. 2 Divergence analysis on the number and length of Aegiceras corniculatum embryos roots in submergence treatments
处理方式 7 d 14 d 21 d 28 d 34 d 胚根长度/cm B 0.33 ± 0.44b 1.09 ± 1.03b 1.60 ± 1.38a 2.02 ± 1.77a 2.45 ± 2.26a C 0.11 ± 0.30a 0.63 ± 0.72a 1.26 ± 0.97a 1.76 ± 1.22a 2.10 ± 1.40a D 0.06 ± 0.16a 0.52 ± 0.51a 1.14 ± 0.69a 1.55 ± 0.83a 2.05 ± 1.10a 胚根数量 B 0.50 ± 0.58b 1.38 ± 1.02a 1.77 ± 1.07a 2.77 ± 1.70a 3.31 ± 1.85a C 0.17 ± 0.38a 0.92 ± 0.97a 1.58 ± 1.02a 2.17 ± 1.05a 2.50 ± 1.06a D 0.15 ± 0.37a 1.04 ± 1.18a 2.65 ± 1.16b 3.69 ± 1.74b 5.04 ± 2.54b 注:不同小写字母表示不同处理组在相同时间下差异显著(p < 0.05)。
下载: 导出CSV
表 3 盐度处理桐花树胚根长度及数量差异性分析
Tab. 3 Divergence analysis on the number and length of Aegiceras corniculatum embryonic roots in salinity treatments
处理方式 7 d 14 d 21 d 28 d 34 d 胚根长度/cm E 0.04 ± 0.14a 0.62 ± 0.51b 1.23 ± 0.96c 1.76 ± 1.34c 2.19 ± 1.63c F 0.17 ± 0.27b 0.54 ± 0.57b 0.81 ± 0.64b 1.09 ± 0.81b 1.24 ± 0.94b G 0.01 ± 0.05a 0.01 ± 0.06a 0.01 ± 0.06a 0.07 ± 0.18a 0.07 ± 0.19a 胚根数量 E 0.07 ± 0.25a 1.10 ± 1.09b 1.53 ± 1.17b 2.40 ± 1.89b 2.93 ± 2.42b F 0.37 ± 0.56b 1.00 ± 1.11b 2.00 ± 1.23b 2.60 ± 1.54b 3.07 ± 1.82b G 0.03 ± 0.18a 0.03 ± 0.18a 0.03 ± 0.18a 0.43 ± 1.30a 0.43 ± 1.30a 注:不同小写字母表示不同处理组在相同时间下差异显著(p < 0.05)。
下载: 导出CSV
表 4 淹水时长−盐度交互处理下胚胎萌根状态
Tab. 4 Embryonic rooting status under the interaction of submergence duration and salinity
处理组 始萌根日数/d 平均萌根日数/d 萌根率/% 平均萌根率/% BE 6.0 11.9 90.0 70.0 CE 4.0 9.9 90.0 50.0 DE 8.0 11.1 80.0 50.0 BF 1.0 8.0 90.0 60.0 CF 5.0 12.0 80.0 40.0 DF 7.0 14.4 100.0 50.0 BG − − 0.0 0.0 CG 6.0 6.0 10.0 10.0 DG 23.0 24.7 30.0 20.0 注:BE处理组为完全浸泡胚胎6 h/d + 浸泡胚根18 h/d,盐度为0;CE处理组为浸泡胚根,盐度为0;DE处理组为蛭石浅插,盐度为0;BF处理组为完全浸泡胚胎6 h/d + 浸泡胚根18 h/d,盐度为10;CF处理组为处理组为浸泡胚根,盐度为10;DF处理组为蛭石浅插,盐度为10;BG处理组为完全浸泡胚胎6 h/d + 浸泡胚根18 h/d,盐度为20;CG处理组为浸泡胚根,盐度为20;DG处理组为蛭石浅插,盐度为20。“−”代胚根无萌动。
下载: 导出CSV
表 5 淹水时长−盐度对胚根长度特征的双因素方差分析
Tab. 5 A bivariate anove of the effects of submergence duration and salinity on the characteristics of radical length
差异来源 7 d时胚根长度 14 d时胚根长度 21 d时胚根长度 28 d时胚根长度 34 d时胚根长度 F p F p F p F p F p 淹水−盐度处理 2.556 0.024 2.749 0.016 4.972 < 0.001 5.496 < 0.001 5.465 < 0.001
下载: 导出CSV
表 6 淹水时长−盐度对胚根数量特征的双因素方差分析
Tab. 6 Bivariate anova of the effects of submergence duration and salinity on the quantitative characteristics of embryonic roots
差异来源 7 d时胚根数量 14 d时胚根数量 21 d时胚根数量 28 d时胚根数量 34 d时胚根数量 F p F p F p F p F p 淹水−盐度处理 2.566 0.023 1.500 0.185 3.914 < 0.001 2.127 0.056 2.404 0.032
下载: 导出CSV
表 7 淹水时长−盐度处理下桐花树胚根生长信息主成分分析
Tab. 7 Principal component analysis of root growth information of Aegiceras corniculatum under the condition of submergence duration and salinity treatment
项目 第一主成分 第二主成分 特征根 3.87 1.66 方差贡献率/% 64.52 27.62 累积方差贡献率/% 64.52 92.14
下载: 导出CSV
表 8 淹水时长−盐度处理下桐花树胚根生长指标的主成分得分
Tab. 8 Principal component scores of root growth indices of Aegiceras corniculatum under the condition of submergence duration and salinity treatment
处理 第一主成分(F1) 第二主成分(F2) 综合得分(F) AE −1.98 0.13 −1.24 AF −1.98 0.13 −1.24 AG −1.98 0.13 −1.24 BE 1.57 −1.13 0.70 BF 2.71 3.48 2.71 BG −1.98 0.13 −1.24 CE 2.01 −1.43 0.91 CF 1.27 0.03 0.83 CG −1.57 0.58 −0.85 DE 1.26 −1.44 0.41 DF 2.23 −0.54 1.29 DG −1.58 −0.08 −1.04
下载: 导出CSV
-
[1] 李春干, 代华兵. 1960−2010年广西红树林空间分布演变机制[J]. 生态学报, 2015, 35(18): 5992−6006.Li Chungan, Dai Huabing. Mechanism analysis of temporal dynamics in mangrove spatial distribution in Guangxi, China: 1960−2010[J]. Acta Ecologica Sinica, 2015, 35(18): 5992−6006. [2] 张忠华, 胡刚, 梁士楚. 广西红树林资源与保护[J]. 海洋环境科学, 2007, 26(3): 275−279, 282. doi: 10.3969/j.issn.1007-6336.2007.03.018Zhang Zhonghua, Hu Gang, Liang Shichu. Mangrove resources and conservation in Guangxi[J]. Marine Environmental Science, 2007, 26(3): 275−279, 282. doi: 10.3969/j.issn.1007-6336.2007.03.018 [3] 范航清, 莫竹承. 广西红树林恢复历史、成效及经验教训[J]. 广西科学, 2018, 25(4): 363−371, 387.Fan Hangqing, Mo Zhucheng. The history, achievements and lessons learnt for mangrove restoration in Guangxi, China[J]. Guangxi Sciences, 2018, 25(4): 363−371, 387. [4] 何斌源, 赖廷和. 广西沿海红海榄造林的宜林临界线[J]. 应用生态学报, 2007, 18(8): 1702−1708.He Binyuan, Lai Tinghe. Critical tidal level for forestation with hypocotyls of Rhizophora stylosa griff along Guangxi coast of China[J]. Chinese Journal of Applied Ecology, 2007, 18(8): 1702−1708. [5] 张乔民, 隋淑珍, 张叶春, 等. 红树林宜林海洋环境指标研究[J]. 生态学报, 2001, 21(9): 1427−1437.Zhang Qiaomin, Sui Shuzhen, Zhang Yechun, et al. Marine environmental indexes related to mangrove growth[J]. Acta Ecologica Sinica, 2001, 21(9): 1427−1437. [6] 刘双双, 杨升, 刘星, 等. 红树林对潮汐浸淹适应性研究进展[J]. 世界林业研究, 2022, 35(5): 25−30.Liu Shuangshuang, Yang Sheng, Liu Xing, et al. Research progress in the adaptability of mangroves to tidal inundation[J]. World Forestry Research, 2022, 35(5): 25−30. [7] 苏柏予, 张维仕, 王友绍. 三种红树根部组织抗氧化酶活性对水淹胁迫的响应[J]. 热带海洋学报, 2022, 41(6): 35−43.Su Boyu, Zhang Weishi, Wang Youshao. Response of antioxidant enzyme systems in root tissues of three mangrove species to waterlogging stress[J]. Journal of Tropical Oceanography, 2022, 41(6): 35−43. [8] 江鎞倩, 李瑞利, 沈小雪, 等. 红树植物耐盐–耐淹性的荟萃分析及其应用对策[J]. 北京大学学报(自然科学版), 2022, 58(4): 687−699.Jiang Biqian, Li Ruili, Shen Xiaoxue, et al. Meta-analysis of mangrove salt-waterlogging tolerance and application strategies[J]. Acta Scientiarum Naturalium Universitatis Pekinensis, 2022, 58(4): 687−699. [9] Cheng Hao, Wu Meilin, Li Changda, et al. Dynamics of radial oxygen loss in mangroves subjected to waterlogging[J]. Ecotoxicology, 2020, 29(6): 684−690. doi: 10.1007/s10646-020-02221-4 [10] 叶功富, 范少辉, 刘荣成, 等. 泉州湾红树林湿地人工生态恢复的研究[J]. 湿地科学, 2005, 1(1): 8−12.Ye Gongfu, Fan Shaohui, Liu Rongcheng, et al. Study on the artificial ecological restoration of mangrove wetlands in Quanzhou Bay[J]. Wetland Science, 2005, 1(1): 8−12. [11] 刁俊明, 刘惠娜, 曾宪录, 等. 不同淡水水位对桐花树幼苗生长的影响[J]. 华南师范大学学报(自然科学版), 2014, 46(2): 108−113.Diao Junming, Liu Huina, Zeng Xianlu, et al. Effects of different fresh water-logging level on the growth and physiological characteristics of Aegiceras corniculatum seedlings[J]. Journal of South China Normal University (Natural Science Edition), 2014, 46(2): 108−113. [12] 郑春芳, 冀德伟, 刘伟成, 等. NaCl胁迫下高纬度移植桐花树幼苗的生理生态效应[J]. 应用生态学报, 2011, 22(9): 2279−2284.Zheng Chunfang, Ji Dewei, Liu Weicheng, et al. Eco-physiological responses of high-latitude transplanted Aegiceras corniculatum seedlings to NaCl stress[J]. Chinese Journal of Applied Ecology, 2011, 22(9): 2279−2284. [13] Solangi K A, Wu Yanyou, Chen Qian, et al. The differential responses of Aegiceras corniculatum and Kandelia candel under salt stress and re-watering phase. A study of leaf electrophysiological and growth parameters[J]. Journal of Plant Interactions, 2021, 16(1): 307−320. doi: 10.1080/17429145.2021.1946606 [14] Ke Lin, Zhang Chunguang, Guo Chuling, et al. Effects of environmental stresses on the responses of mangrove plants to spent lubricating oil[J]. Marine Pollution Bulletin, 2011, 63(5/12): 385−395. [15] 张宜辉. 几种红树植物繁殖体发育和幼苗成长过程的生理生态学研究[D]. 厦门: 厦门大学, 2003.Zhang Yihui. The study of propagule development and seedling growth in some mangrove species[D]. Xiamen: Xiamen University, 2003. [16] 叶勇, 卢昌义, 胡宏友, 等. 三种泌盐红树植物对盐胁迫的耐受性比较[J]. 生态学报, 2004, 24(11): 2444−2450. doi: 10.3321/j.issn:1000-0933.2004.11.014Ye Yong, Lu Changyi, Hu Hongyou, et al. Comparisons of tolerances to salt stress among three salt-secreting mangrove species[J]. Acta Ecologica Sinica, 2004, 24(11): 2444−2450. doi: 10.3321/j.issn:1000-0933.2004.11.014 [17] 陈瑶瑶, 张雅松, 娄铎, 等. 广东英罗湾不同潮位红树林−滩涂系统碳密度差异[J]. 生态环境学报, 2019, 28(6): 1134−1140.Chen Yaoyao, Zhang Yasong, Lou Duo, et al. Carbon density difference of mangrove-shoal system at different tidal levels in Yingluo Bay, Guangdong[J]. Ecology and Environmental Sciences, 2019, 28(6): 1134−1140. [18] 莫竹承, 范航清, 何斌源. 海水盐度对两种红树植物胚轴萌发的影响[J]. 植物生态学报, 2001, 25(2): 235−239. doi: 10.3321/j.issn:1005-264X.2001.02.015Mo Zhucheng, Fan Hangqing, He Binyuan. Effects of seawater salinity on hypocotyl growth in two mangrove species[J]. Chinese Journal of Plant Ecology, 2001, 25(2): 235−239. doi: 10.3321/j.issn:1005-264X.2001.02.015 [19] Jackson M B, Ram P C. Physiological and molecular basis of susceptibility and tolerance of rice plants to complete submergence[J]. Annals of Botany, 2003, 91(2): 227−241. doi: 10.1093/aob/mcf242 [20] 罗美娟, 崔丽娟, 张守攻, 等. 淹水胁迫对桐花树幼苗水分和矿质元素的影响[J]. 福建林学院学报, 2012, 32(4): 336−340.Luo Meijuan, Cui Lijuan, Zhang Shougong, et al. Effects of flooding stress on water and mineral nutrients in Aegiceras corniculatum seedlings[J]. Journal of Fujian College of Forestry, 2012, 32(4): 336−340. [21] 唐莹莹, 郭传超, 石荡, 等. 果肉和埋土深度对新疆野杏种子萌发与幼苗生长的影响[J]. 植物研究, 2023, 43(2): 251−260.Tang Yingying, Guo Chuanchao, Shi Dang, et al. Effects of pulp and buried depth on seed germination and seedling growth of Armeniaca vulgaris[J]. Bulletin of Botanical Research, 2023, 43(2): 251−260. [22] 扈惠灵, 曹永庆, 卫永乐, 等. 磨盘柿杂种胚挽救中的生根壮苗培养[J]. 华北农学报, 2007, 22(5): 117−119.Hu Huiling, Cao Yongqing, Wei Yongle, et al. Rooting and hardening-off culture of Mopanshi persimmon hybrid zygotic embryo[J]. Acta Agriculturae Boreali-Sinica, 2007, 22(5): 117−119. [23] 陈月琴, 蓝崇钰, 黄玉山, 等. 秋茄木榄繁殖体的结构及其生态特异性[J]. 中山大学学报(自然科学版), 1995, 34(4): 70−75.Chen Yueqin, Lan Chongyu, Huang Yushan, et al. Anatomical structures and ecological adaptations of mangrove propagules[J]. Acta Scientiarum Naturalium Universitatis Sunyatseni, 1995, 34(4): 70−75. [24] Zheng Wenjiao, Wang Wenqing, Lin Peng. Dynamics of element contents during the development of hypocotyles and leaves of certain mangrove species[J]. Journal of Experimental Marine Biology and Ecology, 1999, 233(2): 247−257. doi: 10.1016/S0022-0981(98)00131-2 [25] 闫中正, 王文卿, 黄伟滨. 红树胎生现象及其对潮间带生境适应性研究进展[J]. 生态学报, 2004, 24(10): 2317−2323.Yan Zhongzheng, Wang Wenqing, Huang Weibin. Development of the viviparous hypocotyl of mangrove and its adaptation to inter-tidal habitats: a review[J]. Acta Ecologica Sinica, 2004, 24(10): 2317−2323. [26] Wang Wenqing, Yan Zhongzheng, You Siyang, et al. Mangroves: obligate or facultative halophytes? A review[J]. Trees, 2011, 25(6): 953−963. doi: 10.1007/s00468-011-0570-x [27] 张典, 王玉玉, 俞炜炜, 等. 厦门湾红树林生境适宜性评估及修复潜力分析[J]. 应用海洋学学报, 2021, 40(1): 43−55.Zhang Dian, Wang Yuyu, Yu Weiwei, et al. Assessment of the habitat suitability and restoration potential of mangroves in Xiamen Bay, China[J]. Journal of Applied Oceanography, 2021, 40(1): 43−55. [28] 姜仲茂, 管伟, 丁功桃, 等. 不同光照和淹浸程度对木榄幼苗生长的综合效应[J]. 生态环境学报, 2018, 27(10): 1883−1889. doi: 10.16258/j.cnki.1674-5906.2018.10.013Jiang Zhongmao, Guan Wei, Ding Gongtao, et al. Combined effect of different light and inundation on Bruguiera gymnorrhiza seedlings growth[J]. Ecology and Environmental Sciences, 2018, 27(10): 1883−1889. doi: 10.16258/j.cnki.1674-5906.2018.10.013 -

计量
- 文章访问数: 419
- HTML全文浏览量: 192
- PDF下载量: 38
- 被引次数: 0
