

Coral lipids are an important energy source when their symbiotic zooxanthellae density decreases
-
摘要: 近年来珊瑚白化现象日益严峻。白化意味着珊瑚共生虫黄藻密度的降低,然而目前关于珊瑚体内的脂质在虫黄藻密度降低时对维持能量供给稳定的意义尚不清楚。本研究以2020年3月和6月在西沙群岛采集的帛琉蜂巢珊瑚(Favia palauensis)和澄黄滨珊瑚(Porites lutea)样品为材料,通过生理参数(虫黄藻密度、脂质含量)与地球化学指标(虫黄藻的稳定氮同位素δ15Nz值)相结合的方法,探讨了珊瑚脂质对虫黄藻密度及其光合作用强度变化的响应。结果显示,夏季两种珊瑚的虫黄藻密度和δ15Nz值均显著下降,意味着夏季虫黄藻密度降低导致了其光合作用强度的降低。与此同时,两种珊瑚的脂质含量也显著下降,并且脂质含量与虫黄藻密度、δ15Nz值之间均呈正相关关系,这说明珊瑚脂质含量与虫黄藻光合作用强度的变化之间存在耦合关系。当光合作用强度降低时,珊瑚可以通过消耗其自身储存的脂质更好地维持能量供给的稳定,这对提高环境胁迫的适应能力以及抗白化能力具有重要意义。Abstract: Coral bleaching has become increasingly severe in recent years. Bleaching means a decrease in the symbiotic zooxanthellae density in corals. However, the significance of lipids in corals to maintain the stability of energy supply when the zooxanthellae density decreases remains unclear. Favia palauensis and Porites lutea samples collected in the Xisha Islands in March and June 2020 as materials were used in this study. By combining physiological parameters (zooxanthellae density and lipid content) with geochemical indexes (stable nitrogen isotope δ15Nz value of zooxanthellae), the response of coral lipids to the changes in zooxanthellae density and photosynthetic intensity was investigated. The results showed that the zooxanthellae density and δ15Nz value of two genera of corals decreased significantly in summer, implying that the decrease in zooxanthellae density in summer resulted in a decrease in their photosynthetic intensity. At the same time, the lipid content of the two genera of corals also decreased significantly, and the lipid content was positively correlated with zooxanthellae density and δ15Nz value, indicating that there was a coupling relationship between the coral lipid content and the changes of the photosynthetic intensity of zooxanthellae. When photosynthesis intensity decreases, coral can better maintain the stability of energy supply by consuming their own stored lipids, which is of great significance to improve their adaptability to environmental stress and bleaching resilience.
-
Key words:
- coral /
- energy supply /
- zooxanthellae /
- lipids /
- nitrogen isotopes
1) ① 审图号为GS(2020)4618号。 -
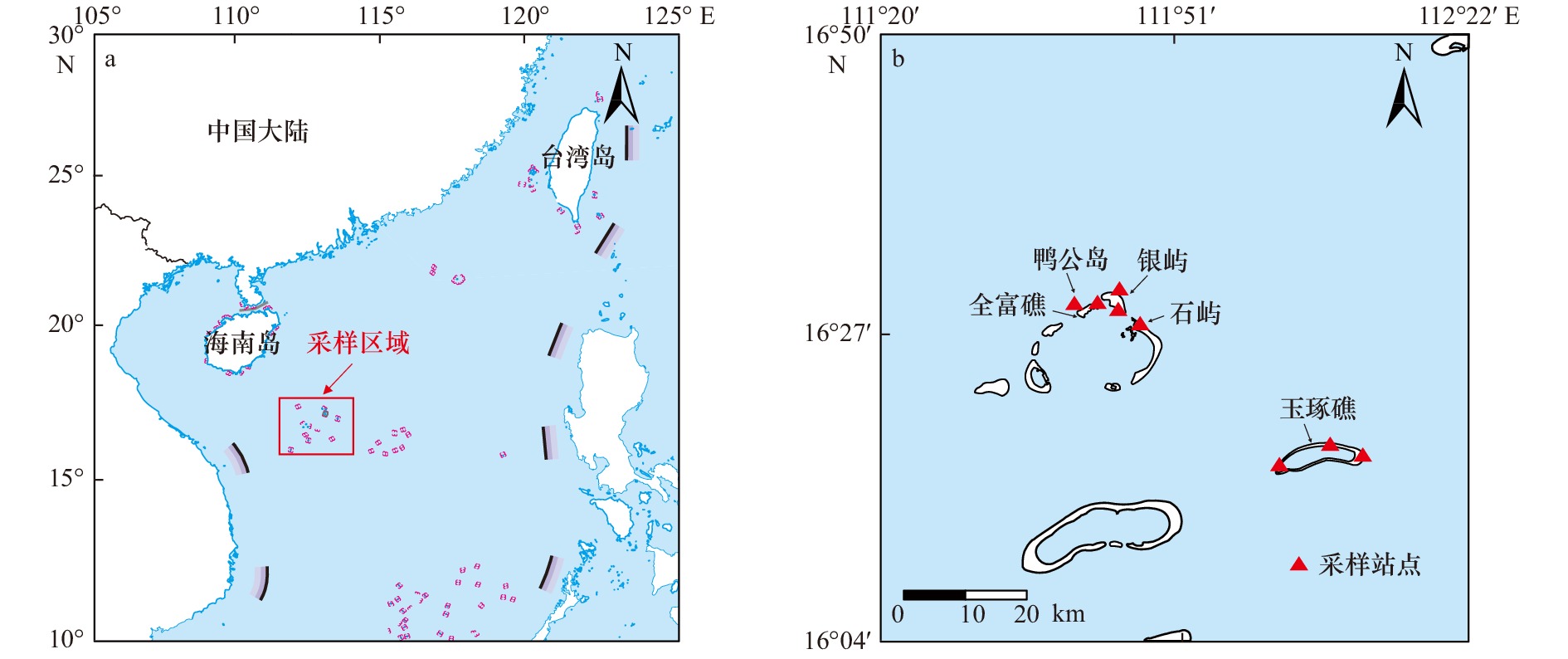
图 1 本文研究区域①(a)和具体采样点(b)
Fig. 1 The study area (a) and specific locations of sampling (b) in this paper
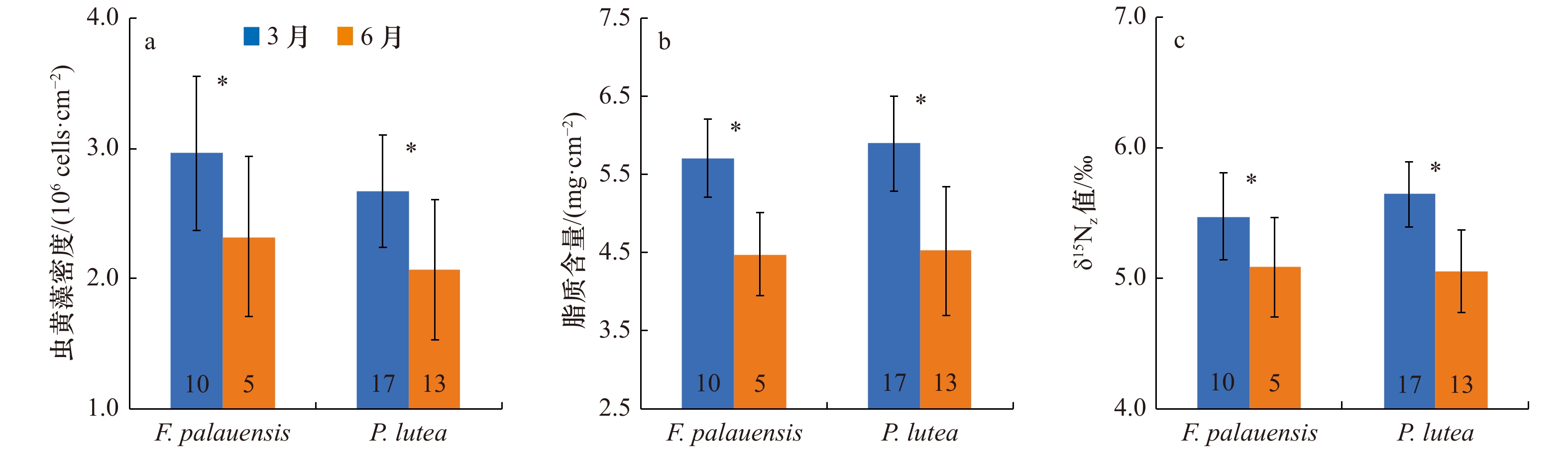
图 2 两种珊瑚虫黄藻密度(a)、脂质含量(b)、δ15Nz值(c)的季节差异
每个条形图上的数字表示珊瑚样本数量;星号表示两种珊瑚每个参数季节间的差异显著(p<0.05)
Fig. 2 Seasonal differences in zooxanthellae density (a), lipid content (b) and δ15Nz value (c) of two genera of corals
The number on each bar chart represents the number of coral samples; the asterisks indicate significant seasonal differences in each parameter of two genera of corals (p<0.05)
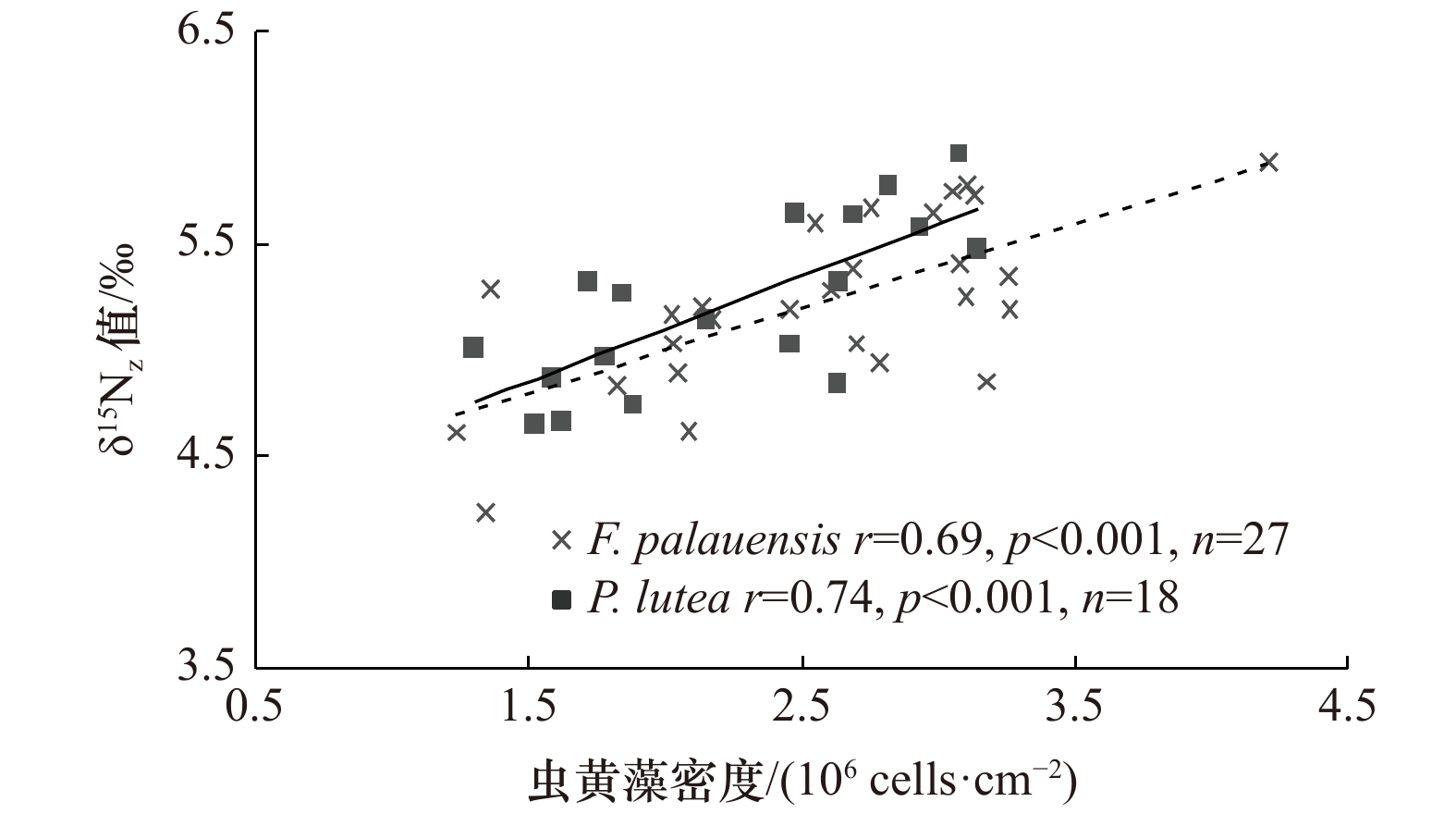
图 3 两种珊瑚虫黄藻密度与δ15Nz值的关系
Fig. 3 Relationship between zooxanthellae density and δ15Nz value of two genera of corals
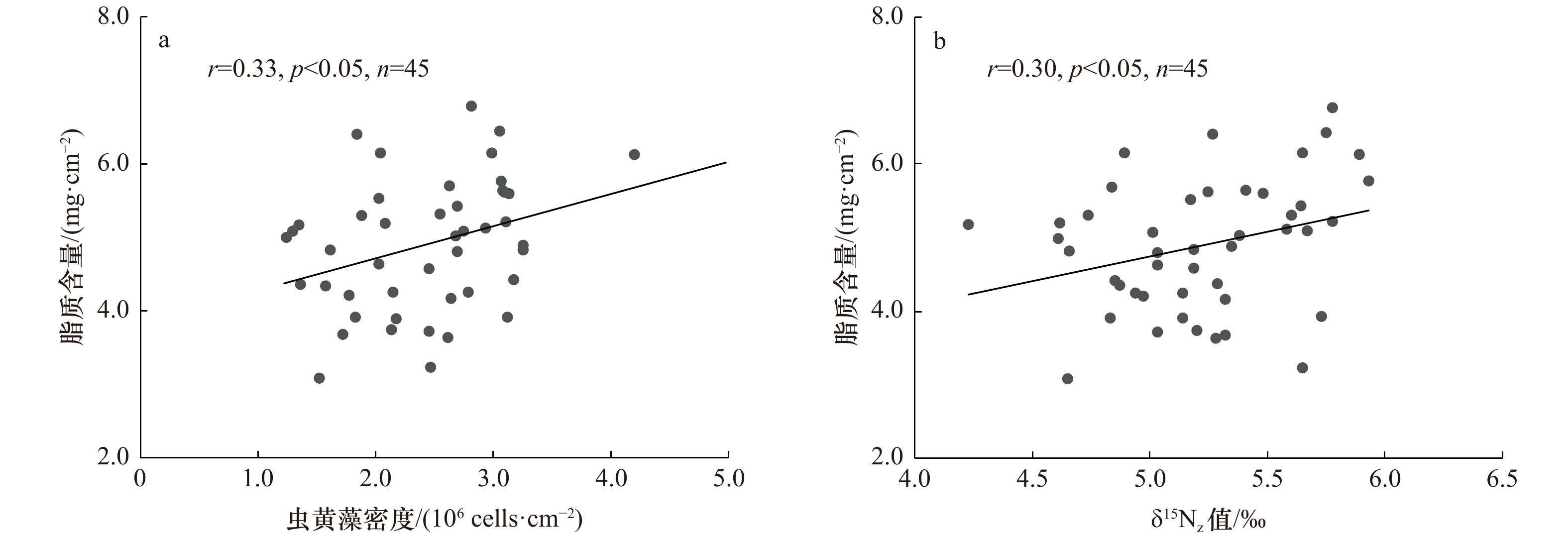
图 4 珊瑚样本虫黄藻密度与脂质含量的关系(a)及珊瑚样本δ15Nz值与脂质含量的关系(b)
Fig. 4 Relationship between zooxanthellae density and lipid content of coral samples (a) and relationship between δ15Nz value and lipid content of coral samples (b)
表 1 两种珊瑚在3月和6月虫黄藻密度、脂质含量、δ15Nz值的属种差异
Tab. 1 Interspecific differences in zooxanthellae density, lipid content and δ15Nz value of two genera of corals in March and June
月份 样品数量 参数 差异值 3月 15 虫黄藻密度 t=0.91 p=0.38 脂质含量 t=−0.69 p=0.50 δ15Nz值 t=−1.04 p=0.32 6月 30 虫黄藻密度 t=1.17 p=0.25 脂质含量 t=0.45 p=0.65 δ15Nz值 t=0.19 p=0.85  下载: 导出CSV
下载: 导出CSV
表 2 两种珊瑚虫黄藻密度、脂质含量、δ15Nz的季节差异
Tab. 2 Seasonal differences in zooxanthellae density, lipid content and δ15Nz of two genera of corals
参数 属种 样品数量 差异值 虫黄藻密度 F. palauensis 27 t=2.53 p<0.05 P. lutea 18 t=2.13 p<0.05 脂质含量 F. palauensis 27 t=5.39 p<0.001 P. lutea 18 t=3.50 p<0.05 δ15Nz值 F. palauensis 27 t=2.71 p<0.05 P. lutea 18 t=3.73 p<0.05
下载: 导出CSV
-
[1] Baumann J, Grottoli A G, Hughes A D, et al. Photoautotrophic and heterotrophic carbon in bleached and non-bleached coral lipid acquisition and storage[J]. Journal of Experimental Marine Biology and Ecology, 2014, 461: 469−478. doi: 10.1016/j.jembe.2014.09.017 [2] Baker A C, Glynn P W, Riegl B. Climate change and coral reef bleaching: an ecological assessment of long-term impacts, recovery trends and future outlook[J]. Estuarine, Coastal and Shelf Science, 2008, 80(4): 435−471. doi: 10.1016/j.ecss.2008.09.003 [3] 吴钟解, 王道儒, 涂志刚, 等. 西沙生态监控区造礁石珊瑚退化原因分析[J]. 海洋学报, 2011, 33(4): 140−146.Wu Zhongjie, Wang Daoru, Tu Zhigang, et al. The analysis on the reason of hermatypic coral degradation in Xisha[J]. Haiyang Xuebao, 2011, 33(4): 140−146. [4] Warner M, Chilcoat G, McFarland F, et al. Seasonal fluctuations in the photosynthetic capacity of photosystem II in symbiotic dinoflagellates in the Caribbean reef-building coral Montastraea[J]. Marine Biology, 2002, 141(1): 31−38. doi: 10.1007/s00227-002-0807-8 [5] 梁甲元, 邓传奇, 许勇前, 等. 一种环境敏感型造礁石珊瑚Pocillopora sp. 共生虫黄藻和细菌的生态特征[J]. 海洋学报, 2022, 44(2): 102−112.Liang Jiayuan, Deng Chuanqi, Xu Yongqian, et al. Ecological characteristics of symbiotic Symbiodiniaceae and bacteria in an environmentally sensitive reef-building coral Pocillopora sp.[J]. Haiyang Xuebao, 2022, 44(2): 102−112. [6] Yamashiro H, Oku H, Onaga K. Effect of bleaching on lipid content and composition of Okinawan corals[J]. Fisheries Science, 2005, 71(2): 448−453. doi: 10.1111/j.1444-2906.2005.00983.x [7] Pengsakun S, Yeemin T, Sutthacheep M, et al. Monitoring of coral communities in the inner Gulf of Thailand influenced by the elevated seawater temperature and flooding[J]. Acta Oceanologica Sinica, 2019, 38(1): 102−111. doi: 10.1007/s13131-019-1376-8 [8] Fitt W K, McFarland F K, Warner M E, et al. Seasonal patterns of tissue biomass and densities of symbiotic dinoflagellates in reef corals and relation to coral bleaching[J]. Limnology and Oceanography, 2000, 45(3): 677−685. doi: 10.4319/lo.2000.45.3.0677 [9] Porter J W, Fitt W K, Spero H J, et al. Bleaching in reef corals: physiological and stable isotopic responses[J]. Proceedings of the National Academy of Sciences of the United States of America, 1989, 86(23): 9342−9346. doi: 10.1073/pnas.86.23.9342 [10] Rodrigues L J, Grottoli A G. Energy reserves and metabolism as indicators of coral recovery from bleaching[J]. Limnology and Oceanography, 2007, 52(5): 1874−1882. doi: 10.4319/lo.2007.52.5.1874 [11] Bessell-Browne P, Stat M, Thomson D, et al. Coscinaraea marshae corals that have survived prolonged bleaching exhibit signs of increased heterotrophic feeding[J]. Coral Reefs, 2014, 33(3): 795−804. doi: 10.1007/s00338-014-1156-z [12] Grottoli A G, Rodrigues L J, Palardy J E. Heterotrophic plasticity and resilience in bleached corals[J]. Nature, 2006, 440(7088): 1186−1189. doi: 10.1038/nature04565 [13] 许慎栋, 张志楠, 余克服, 等. 南海造礁珊瑚Favia palauensis营养方式的空间差异及其对环境适应性的影响[J]. 中国科学: 地球科学, 2021, 64(6): 839−852. doi: 10.1007/s11430-020-9774-0Xu Shendong, Zhang Zhinan, Yu Kefu, et al. Spatial variations in the trophic status of Favia palauensis corals in the South China Sea: insights into their different adaptabilities under contrasting environmental conditions[J]. Science China: Earth Sciences, 2021, 64(6): 839−852. doi: 10.1007/s11430-020-9774-0 [14] Muscatine L. Glycerol excretion by symbiotic algae from corals and Tridacna and its control by the host[J]. Science, 1967, 156(3774): 516−519. doi: 10.1126/science.156.3774.516 [15] Patton J S, Battey J F, Rigler M W, et al. A comparison of the metabolism of bicarbonate 14C and acetate 1-14C and the variability of species lipid compositions in reef corals[J]. Marine Biology, 1983, 75(2): 121−130. [16] Crossland C J, Barnes D J, Borowitzka M A. Diurnal lipid and mucus production in the staghorn coral Acropora acuminata[J]. Marine Biology, 1980, 60(2): 81−90. [17] Patton J S, Abraham S, Benson A A. Lipogenesis in the intact coral Pocillopora capitata and its isolated zooxanthellae: evidence for a light-driven carbon cycle between symbiont and host[J]. Marine Biology, 1977, 44(3): 235−247. doi: 10.1007/BF00387705 [18] Imbs A B, Latyshev N A, Dautova T N, et al. Distribution of lipids and fatty acids in corals by their taxonomic position and presence of zooxanthellae[J]. Marine Ecology Progress Series, 2010, 409: 65−75. doi: 10.3354/meps08622 [19] Yamashiro H, Oku H, Higa H, et al. Composition of lipids, fatty acids and sterols in Okinawan corals[J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 1999, 122(4): 397−407. doi: 10.1016/S0305-0491(99)00014-0 [20] Harland A D, Navarro J C, Davies P S, et al. Lipids of some Caribbean and Red Sea corals: total lipid, wax esters, triglycerides and fatty acids[J]. Marine Biology, 1993, 117(1): 113−117. doi: 10.1007/BF00346432 [21] Grottoli A G, Rodrigues L J, Juarez C. Lipids and stable carbon isotopes in two species of Hawaiian corals, Porites compressa and Montipora verrucosa, following a bleaching event[J]. Marine Biology, 2004, 145(3): 621−631. [22] Stimson J. The annual cycle of density of zooxanthellae in the tissues of field and laboratory-held Pocillopora damicornis (Linnaeus)[J]. Journal of Experimental Marine Biology and Ecology, 1997, 214(1/2): 35−48. [23] 李淑, 余克服, 施祺, 等. 南海北部珊瑚共生虫黄藻密度的种间与空间差异及其对珊瑚礁白化的影响[J]. 科学通报, 2008, 53(2): 295−303. doi: 10.1007/s11434-007-0514-4Li Shu, Yu Kefu, Shi Qi, et al. Interspecies and spatial diversity in the symbiotic zooxanthellae density in corals from northern South China Sea and its relationship to coral reef bleaching[J]. Chinese Science Bulletin, 2008, 53(2): 295−303. doi: 10.1007/s11434-007-0514-4 [24] Grottoli A G, Rodrigues L J. Bleached Porites compressa and Montipora capitata corals catabolize δ13C-enriched lipids[J]. Coral Reefs, 2011, 30(3): 687−692. doi: 10.1007/s00338-011-0756-0 [25] Oku H, Yamashiro H, Onaga K. Lipid biosynthesis from [14C]-glucose in the coral Montipora digitata[J]. Fisheries Science, 2003, 69(3): 625−631. doi: 10.1046/j.1444-2906.2003.00665.x [26] Heikoop J M, Risk M J, Lazier A V, et al. Nitrogen-15 signals of anthropogenic nutrient loading in reef corals[J]. Marine Pollution Bulletin, 2000, 40(7): 628−636. doi: 10.1016/S0025-326X(00)00006-0 [27] Muscatine L, Kaplan I R. Resource partitioning by reef corals as determined from stable isotope composition II. δ15N of zooxanthellae and animal tissue versus depth[J]. Pacific Science, 1994, 48(3): 304−312. [28] Ferrier-Pagès C, Peirano A, Abbate M, et al. Summer autotrophy and winter heterotrophy in the temperate symbiotic coral Cladocora caespitosa[J]. Limnology and Oceanography, 2011, 56(4): 1429−1438. doi: 10.4319/lo.2011.56.4.1429 [29] Heikoop J M, Dunn J J, Risk M J, et al. Relationship between light and the δ15N of coral tissue: examples from Jamaica and Zanzibar[J]. Limnology and Oceanography, 1998, 43(5): 909−920. doi: 10.4319/lo.1998.43.5.0909 [30] Djeghri N, Stibor H, Lebeau O, et al. δ13C, δ15N, and C∶N ratios as nutrition indicators of zooxanthellate jellyfishes: insights from an experimental approach[J]. Journal of Experimental Marine Biology and Ecology, 2020, 522: 151257. doi: 10.1016/j.jembe.2019.151257 [31] Qin Zhenjun, Yu Kefu, Wang Yinghui, et al. Spatial and intergeneric variation in physiological indicators of corals in the South China Sea: insights into their current state and their adaptability to environmental stress[J]. Journal of Geophysical Research: Oceans, 2019, 124(5): 3317−3332. doi: 10.1029/2018JC014648 [32] 李颖虹, 黄小平, 岳维忠. 西沙永兴岛环境质量状况及管理对策[J]. 海洋环境科学, 2004, 23(1): 50−53. doi: 10.3969/j.issn.1007-6336.2004.01.015Li Yinghong, Huang Xiaoping, Yue Weizhong. Environmental quality and management measures in Yongxing Island of Xisha, South China Sea[J]. Marine Environmental Science, 2004, 23(1): 50−53. doi: 10.3969/j.issn.1007-6336.2004.01.015 [33] Fagoonee I, Wilson H B, Hassell M P, et al. The dynamics of zooxanthellae populations: a long-term study in the field[J]. Science, 1999, 283(5403): 843−845. doi: 10.1126/science.283.5403.843 [34] Seemann J. The use of 13C and 15N isotope labeling techniques to assess heterotrophy of corals[J]. Journal of Experimental Marine Biology and Ecology, 2013, 442: 88−95. doi: 10.1016/j.jembe.2013.01.004 [35] Stimson J S. Location, quantity and rate of change in quantity of lipids in tissue of Hawaiian Hermatypic corals[J]. Bulletin of Marine Science, 1987, 41(3): 889−904. [36] Rodrigues L J, Grottoli A G. Calcification rate and the stable carbon, oxygen, and nitrogen isotopes in the skeleton, host tissue, and zooxanthellae of bleached and recovering Hawaiian corals[J]. Geochimica et Cosmochimica Acta, 2006, 70(11): 2781−2789. doi: 10.1016/j.gca.2006.02.014 [37] Glynn P W, D'Croz L. Experimental evidence for high temperature stress as the cause of El Niño-coincident coral mortality[J]. Coral Reefs, 1990, 8(4): 181−191. doi: 10.1007/BF00265009 [38] Fujise L, Yamashita H, Suzuki G, et al. Moderate thermal stress causes active and immediate expulsion of photosynthetically damaged zooxanthellae (Symbiodinium) from corals[J]. PLoS One, 2014, 9(12): e114321. doi: 10.1371/journal.pone.0114321 [39] Gates R D, Baghdasarian G, Muscatine L. Temperature stress causes host cell detachment in symbiotic cnidarians: implications for coral bleaching[J]. The Biological Bulletin, 1992, 182(3): 324−332. doi: 10.2307/1542252 [40] Lesser M P. Elevated temperatures and ultraviolet radiation cause oxidative stress and inhibit photosynthesis in ymbiotic dinoflagellates[J]. Limnology and Oceanography, 1996, 41(2): 271−283. doi: 10.4319/lo.1996.41.2.0271 [41] Kuguru B, Winters G, Beer S, et al. Adaptation strategies of the corallimorpharian Rhodactis rhodostoma to irradiance and temperature[J]. Marine Biology, 2007, 151(4): 1287−1298. doi: 10.1007/s00227-006-0589-5 [42] Salih A, Larkum A, Cox G, et al. Fluorescent pigments in corals are photoprotective[J]. Nature, 2000, 408(6814): 850−853. doi: 10.1038/35048564 [43] Brown B E, Downs C A, Dunne R P, et al. Exploring the basis of thermotolerance in the reef coral Goniastrea aspera[J]. Marine Ecology Progress Series, 2002, 242: 119−129. doi: 10.3354/meps242119 [44] Muscatine L, Cernichiari E. Assimilation of photosynthetic products of zooxanthellae by a reef coral[J]. The Biological Bulletin, 1969, 137(3): 506−523. doi: 10.2307/1540172 [45] Davies P S. Effect of daylight variations on the energy budgets of shallow-water corals[J]. Marine Biology, 1991, 108(1): 137−144. doi: 10.1007/BF01313481 [46] Lesser M P. Using energetic budgets to assess the effects of environmental stress on corals: are we measuring the right things?[J]. Coral Reefs, 2013, 32(1): 25−33. doi: 10.1007/s00338-012-0993-x [47] Kellogg R B, Patton J S. Lipid droplets, medium of energy exchange in the symbiotic anemone Condylactis gigantea: a model coral polyp[J]. Marine Biology, 1983, 75(2): 137−149. [48] Anthony K R N, Connolly S R, Willis B L. Comparative analysis of energy allocation to tissue and skeletal growth in corals[J]. Limnology and Oceanography, 2002, 47(5): 1417−1429. doi: 10.4319/lo.2002.47.5.1417 [49] Gnaiger E, Bitterlich G. Proximate biochemical composition and caloric content calculated from elemental CHN analysis: a stoichiometric concept[J]. Oecologia, 1984, 62(3): 289−298. doi: 10.1007/BF00384259 [50] Anthony K R N, Hoogenboom M O, Maynard J A, et al. Energetics approach to predicting mortality risk from environmental stress: a case study of coral bleaching[J]. Functional Ecology, 2009, 23(3): 539−550. doi: 10.1111/j.1365-2435.2008.01531.x -

计量
- 文章访问数: 1064
- HTML全文浏览量: 401
- PDF下载量: 88
- 被引次数: 0
