

The association of single nucleotide polymorphisms of the glycogen phosphorylase gene with glycogen content in the razor clam Sinonovacula constricta
-
摘要: 为探究缢蛏糖原磷酸化酶基因(Sc-GPH)与糖原含量的关系,本研究克隆获得Sc-GPH cDNA全长,检测其在不同组织、不同月份的表达模式,并在Sc-GPH基因编码区筛选与糖原含量关联的SNP位点。结果表明,Sc-GPH cDNA全长为3 963 bp,开放阅读框为2 541 bp,编码846个氨基酸;氨基酸序列多重比对和系统发育树显示,缢蛏与欧洲大扇贝、虾夷扇贝、长牡蛎等贝类亲缘关系较近,而与哺乳类、甲壳类、昆虫类亲缘关系较远。实时荧光定量PCR结果显示,Sc-GPH在8个组织中均有表达,其中在外套膜和足中表达量最高(p<0.01),推测与其糖原存储能力有关;不同月份的缢蛏外套膜和足中,Sc-GPH在8月表达量最高,而此时糖原含量较低,说明Sc-GPH的表达可能受缢蛏生殖周期的影响。以缢蛏“甬乐1号”群体为实验材料,在Sc-GPH基因编码区筛选到4个与糖原含量相关SNP位点,其中c.930T>C位点在台州野生群体中得到进一步验证,该位点为高糖原缢蛏分子标记辅助选育提供了候选标记。Abstract: To clarify the relationship between the glycogen phosphorylase gene (Sc-GPH) and glycogen content in Sinonovacula constricta, the full-length cDNA sequence of Sc-GPH is obtained using RACE technology, its expression pattern in different tissues, different months are analyzed, and the SNP loci of Sc-GPH associated with glycogen content are explored. The full-length cDNA of Sc-GPH gene is 3 963 bp and the ORF is 2 541 bp, encoding 846 amino acids. The multiple alignment of amino acid sequences and phylogenetic tree show that S. constricta are closely related to shellfish such as Pecten maximus, Mizuhopecten yessoens, Crassostrea gigas, but farther to mammals, arthropodas and insects. The results of real-time quantitative PCR show that Sc-GPH gene is expressed in eight tissues, with the highest expression in mantle and foot, suggesting they are related to glycogen storage capacity. In different months, the expression levels of Sc-GPH in the mantle and foot is the highest in August, at which time the glycogen content decreased, indicating that the expression of Sc-GPH seems to be driven by reproductive cycle of S. constricta. In “YL1” population, four SNPs related to glycogen content are screened in the coding region of Sc-GPH gene, and c.930T>C is further verified in Taizhou wild population, which provides a candidate marker for molecular marker assisted breeding of high glycogen S. constricta varieties.
-
Key words:
- Sinonovacula constricta /
- glycogen phosphorylase /
- glycogen content /
- SNP /
- correlation analysis
-
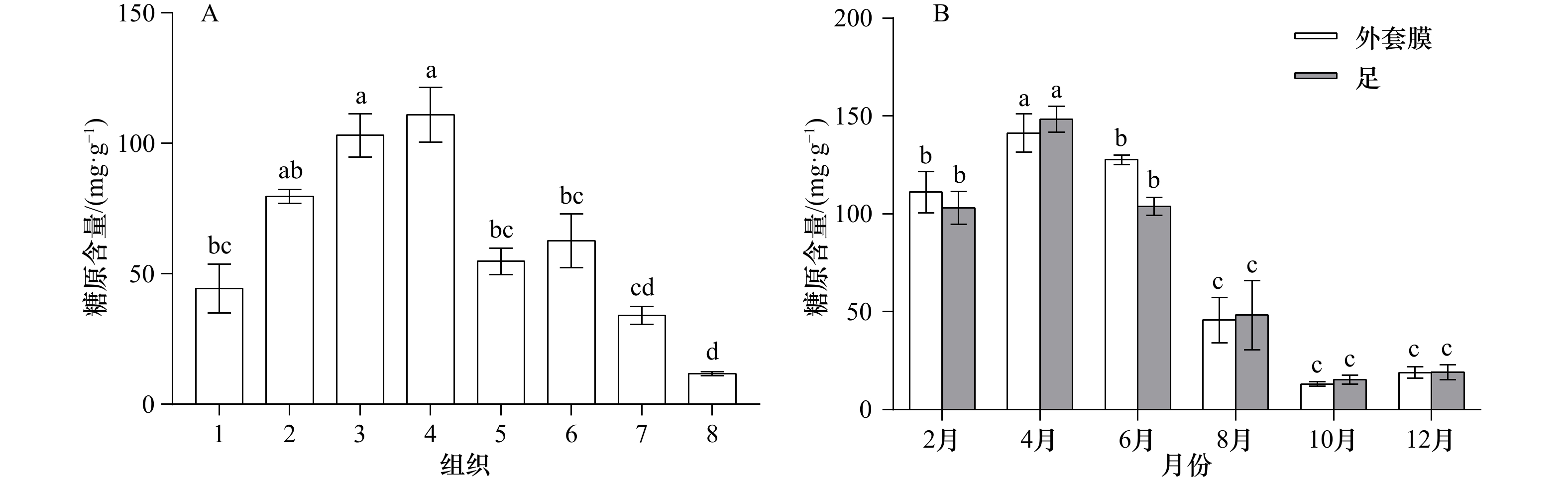
图 1 缢蛏不同组织(A)和不同月份足和外套膜(B)的糖原含量变化
1. 鳃;2. 闭壳肌;3. 足;4. 外套膜;5. 肝胰腺;6. 性腺;7. 水管;8. 唇瓣;A中不同小写字母代表差异极显著(p<0.01);B中同一组织不同字母代表差异极显著(p<0.01)
Fig. 1 Changes of glycogen content in different tissues (A) and different months in foot and mantle (B) of Sinonovacula constricta
1. Gill; 2. adductor muscle; 3. foot; 4. mantle; 5. hepatopancreas; 6. gonad; 7. siphon; 8. palp; different lowercases indicate extremely significant difference (p<0.01) in A; different letters marked on columns of the same tissue represent their extremely significant differences in data (p<0.01) in B

图 2 缢蛏与其他物种GPH氨基酸序列系统进化分析
进化树中各物种GPH氨基酸序列的GenBank登录号为:缢蛏(Sinonovacula constricta,MT125681);长牡蛎(Crassostrea gigas,CCN27372);欧洲大扇贝(Pecten maximus,XP_033734012);虾夷扇贝(Mizuhopecten yessoensis,OWF50424);克氏原螯虾(Procambarus clarki,AVN99053);凡纳滨对虾(Penaeus vannamei,QCY50320);日本对虾(Procambarus clarkii,BAJ23879);异色瓢虫(Harmonia axyridis,ASZ80181);黑腹果蝇(Drosophila melanogaster,NP_001027219);家蚕(Bombyx mori,ACB41088);甜菜夜蛾(Spodoptera exigua,ACN78408);亚洲玉米螟(Ostrinia furnacalis,AFO54708);智人(Homo sapiens,AAC18079);小家鼠(Mus musculus,AAG00588);马(Equus caballus,BAH22533);单峰驼(Camelus dromedarius,KAB1277008)
Fig. 2 Neighbor-joining phylogenetic tree of GPH among Sinonovacula constricta and other species
GenBank accession numbers of GPH sequences used for phylogenetic tree: Razor clam (Sinonovacula constricta, MT125681); Pacific oyster (Crassostrea gigas, CCN27372); European scallop (Pecten maximus, XP_033734012); Japanese scallop (Mizuhopecten yessoensis, OWF50424); Red swamp crayfish (Procambarus clarki, AVN99053); Whiteleg shrimp (Penaeus vannamei, QCY50320); Kuruma prawn (Penaeus japonicus, BAJ23879); Asian lady beetles (Harmonia axyridis, ASZ80181); Fruit fly (Drosophila melanogaster, NP_001027219); Domestic silkworm (Bombyx mori, ACB41088); Beet armyworm (Spodoptera exigua, ACN78408); Asiatic corn borer (Ostrinia furnacalis, AFO54708); Human (Homo sapiens, AAC18079); House mouse (Mus musculus, AAG00588); Horse (Equus caballus, BAH22533); Single hump camel (Camelus dromedarius, KAB1277008)
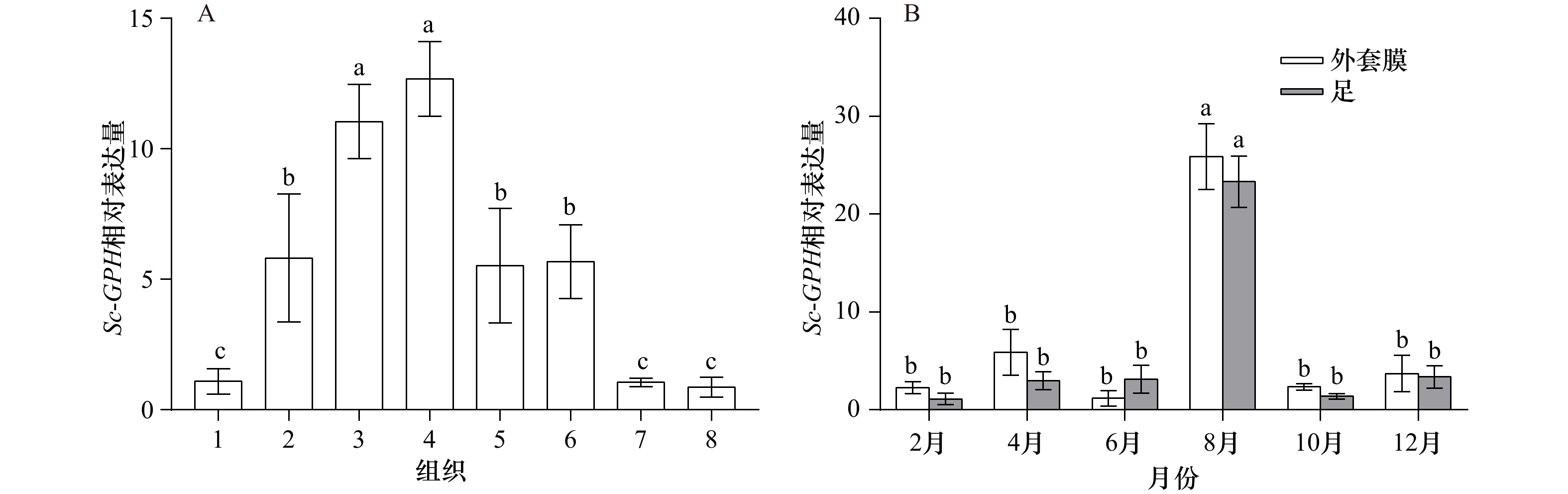
图 3 Sc-GPH在缢蛏不同组织(A)和不同月份足和外套膜(B)的的表达特征
1. 鳃;2. 闭壳肌;3. 足;4. 外套膜;5. 肝胰腺;6. 性腺;7. 水管;8. 唇瓣;A中不同小写字母代表差异极显著(p<0.01);B中同一组织不同字母代表差异极显著(p<0.01)
Fig. 3 The expression characteristics of Sc-GPH in different tissues (A) and different months in foot and mantle (B)
1. Gill; 2. adductor muscle; 3. foot; 4. mantle; 5. hepatopancreas; 6. gonad; 7. siphon; 8. palp; different lowercases indicate extremely significant difference(p<0.01)in A; different letters marked on columns of the same tissue represent their extremely significant differences in data (p<0.01)in B
表 1 所用的引物及信息
Tab. 1 The information of primers used in this experiment
引物名称 引物序列(5′-3′) 用途 H3 AGTGAACAGATCTCCACCGCAGGCA Sc-GPH 3′-RACE扩增 H5 GCGGGATGACTTGAACCTACGGAT Sc-GPH 5′-RACE扩增 18S-F TCGGTTCTATTGCGTTGGTTTT qPCR 18S-R CAGTTGGCATCGTTTATGGTCA qPCR H-Q-F ATCTACAACTCCCTCCTCTACCA qPCR H-Q-R GGCACATCTTAGCCCAAAG qPCR H-S-F AAAGACATTCAATCGCCACCT SNP筛选 H-S-R TTTCTGGATTTGGCTCACAACCC SNP筛选  下载: 导出CSV
下载: 导出CSV
表 2 缢蛏“甬乐1号”(YL1)和台州野生群体(TZ)Sc-GPH SNPs位点与糖原含量的相关性分析
Tab. 2 Association of SNPs of Sc-GPH with glycogen content in YL1 and TZ of Sinonovacula constricta
群体 SNP位点 基因型 样本量/频率 糖原含量/(mg·g−1) p值 YL1 c.874C>T CC 89/0.89 54.6±30.3 0.047* CT 11/0.11 74.1±30.0 c.930T>C TT 49/0.49 52.1±29.4 0.029* TC 43/0.43 57.9±28.4 CC 8/0.08 78.6±34.7 c.1133 T>C TT 87/0.87 53.6±29.2 0.023* TC 13/0.13 77.7±27.1 c.1284 T>A TT 65/0.65 61.0±26.7 0.028* TA 23/0.23 56.8±37.7 AA 12/0.12 33.1±1.97 TZ c.930T>C TT 47/0.47 73.9±22.5 0.047* TC 40/0.4 80.7±23.2 CC 13/0.13 86.1±15.0 注:p值代表SNP位点与糖原含量的相关程度,*代差异显著(p<0.05)。
下载: 导出CSV
-
[1] Greenberg C C, Jurczak M J, Danos A M, et al. Glycogen branches out: new perspectives on the role of glycogen metabolism in the integration of metabolic pathways[J]. American Journal of Physiology-Endocrinology and Metabolism, 2006, 291(1): E1−E8. doi: 10.1152/ajpendo.00652.2005 [2] Berthelin C, Kellner K, Mathieu M. Storage metabolism in the Pacific oyster (Crassostrea gigas) in relation to summer mortalities and reproductive cycle (west coast of France)[J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2000, 125(3): 359−369. doi: 10.1016/S0305-0491(99)00187-X [3] Liu Wenguang, Li Qi, Yuan Yundang, et al. Seasonal variations in reproductive activity and biochemical composition of the cockle Fulvia mutica (Reeve) from the eastern coast of China[J]. Journal of Shellfish Research, 2008, 27(2): 405−411. doi: 10.2983/0730-8000(2008)27[405:SVIRAA]2.0.CO;2 [4] Park M S, Kang C K, Lee P Y. Reproductive cycle and biochemical composition of the ark shell Scapharca broughtonii (Schrenck) in a southern coastal bay of Korea[J]. Journal of Shellfish Research, 2001, 20(1): 177−184. [5] 闫红伟. 缢蛏 (Sinonovacula constricta) 和青蛤 (Cyclina sinensis) 繁殖生理学的研究[D]. 青岛: 中国海洋大学, 2009: 32−33Yan Hongwei. Reproductive physiology of Sinonovacula constricta and Cyclina sinensis [D]. Qingdao: Ocean University of China, 2009: 32−33. [6] Samain J F. Review and perspectives of physiological mechanisms underlying genetically-based resistance of the Pacific oyster Crassostrea gigas to summer mortality[J]. Aquatic Living Resources, 2011, 24(3): 227−236. doi: 10.1051/alr/2011144 [7] 陈德慰, 苏键, 刘小玲, 等. 广西北部湾3种贝类中主要呈味物质的测定及呈味作用评价[J]. 食品科学, 2012, 33(10): 165−168.Chen Dewei, Su Jian, Liu Xiaoling, et al. Taste evaluation of non-volatile taste compounds in bivalve mollusks from Beibu gluf, Guangxi[J]. Food Science, 2012, 33(10): 165−168. [8] 李含. 缢蛏综合保活技术研究[D]. 福州: 福建农林大学, 2014: 37−41Li Han. The research of comprehensive keep-alive technology to Sinonovacula constricta[D]. Fuzhou: Fujian Agriculture and Forestry University, 2014: 37−41. [9] Palsamy P, Subramanian S. Modulatory effects of resveratrol on attenuating the key enzymes activities of carbohydrate metabolism in streptozotocin–nicotinamide-induced diabetic rats[J]. Chemico-Biological Interactions, 2009, 179(2/3): 356−362. [10] Cori C F, Cori G T. Mechanism of formation of hexosemonophosphate in muscle and isolation of a new phosphate ester[J]. Experimental Biology and Medicine, 1936, 34(5): 702−705. doi: 10.3181/00379727-34-8759P [11] Nakano K, Hwang P K, Fletterick R J. Complete cDNA sequence for rabbit muscle glycogen phosphorylase[J]. Federation of European Biochemical Societies, 1986, 204(2): 283−287. doi: 10.1016/0014-5793(86)80829-8 [12] Bacca H, Huvet A, Fabioux C, et al. Molecular cloning and seasonal expression of oyster glycogen phosphorylase and glycogen synthase genes[J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2005, 140(4): 635−646. doi: 10.1016/j.cbpc.2005.01.005 [13] Zeng Zhen, Ni Jianbin, Ke Caihuan. Glycogen content relative to expression of glycogen phosphorylase (GPH) and hexokinase (HK) during the reproductive cycle in the Fujian oyster, Crassostrea angulata[J]. Acta Oceanologica Sinica, 2015, 34(6): 66−76. doi: 10.1007/s13131-015-0639-2 [14] She Zhicai, Li Li, Qi Haigang, et al. Candidate gene polymorphisms and their association with glycogen content in the Pacific oyster Crassostrea gigas[J]. PLoS One, 2015, 10(5): e0124401. doi: 10.1371/journal.pone.0124401 [15] 霍礼辉, 陈彩芳, 林志华, 等. 镉诱导缢蛏(Sinonovacula constricta)体内金属硫蛋白基因变化规律研究[J]. 海洋与湖沼, 2012, 43(4): 723−728. doi: 10.11693/hyhz201204006006Huo Lihui, Chen Caifang, Lin Zhihua, et al. The metallothionein gene expression in Sinonovacula constricta exposing to cadmium[J]. Oceanologia et Limnologia Sinica, 2012, 43(4): 723−728. doi: 10.11693/hyhz201204006006 [16] 孙莎, 钟婵, 章骞, 等. 4种海产贝类中主要呈味物质含量与季节变化的关系[J]. 集美大学学报(自然科学版), 2016, 21(6): 420−427.Sun Sha, Zhong Chan, Zhang Qian, et al. Seasonal variation of taste-active component contents in four species of marine shellfish[J]. Journal of Jimei University (Natural Science), 2016, 21(6): 420−427. [17] 陈燕园, 刘圣, 何京, 等. 浙江缢蛏群体糖原含量变化与gys基因多态性的关联分析[J]. 水产学报, 2021, 45(3): 415−423.Chen Yanyuan, Liu Sheng, He Jing, et al. Glycogen content fluctuation and association analysis with polymorphism of gys gene of Sinonovacula constricta in the Zhejiang population[J]. Journal of Fisheries of China, 2021, 45(3): 415−423. [18] Bayne B L. Aspects of reproduction in bivalve molluscs[M]//Wiley M. Estuarine Processes. New York: Academic Press, 1976: 432−448. [19] Barber B J, Blake N J. Energy storage and utilization in relation to gametogenesis in Argopecten irradians concentricus (say)[J]. Journal of Experimental Marine Biology and Ecology, 1981, 52(2/3): 121−134. [20] Yan Hongwei, Li Qi, Liu Wenguang, et al. Seasonal changes in reproductive activity and biochemical composition of the razor clam Sinonovacula constricta (Lamarck 1818)[J]. Marine Biology Research, 2010, 6(1): 78−88. doi: 10.1080/17451000903039756 [21] Sandström M E, Abbate F, Andersson D C, et al. Insulin-independent glycogen supercompensation in isolated mouse skeletal muscle: role of phosphorylase inactivation[J]. Pflügers Archiv, 2004, 448(5): 533−538. [22] Ferguson D M, Gerrard D E. Regulation of post-mortem glycolysis in ruminant muscle[J]. Animal Production Science, 2014, 54(4): 464−481. doi: 10.1071/AN13088 [23] Zhao Na, Hou Ming, Wang Ting, et al. Cloning and expression patterns of the brine shrimp (Artemia sinica) glycogen phosphorylase (GPase) gene during development and in response to temperature stress[J]. Molecular Biology Reports, 2014, 41(1): 9−18. doi: 10.1007/s11033-013-2764-3 [24] Vali S, Carlsen R, Pessah I, et al. Role of the sarcoplasmic reticulum in regulating the activity-dependent expression of the glycogen phosphorylase gene in contractile skeletal muscle cells[J]. Journal of Cellular Physiology, 2000, 185(2): 184−199. doi: 10.1002/1097-4652(200011)185:2<184::AID-JCP3>3.0.CO;2-T [25] Oruç Ö E, Dursun Ö. An application of information theoretical measures for DNA structure[J]. Turkiye Klinikleri Journal of Biostatistics, 2011, 3(1): 1−7. [26] Fiume E, Christou P, Giani S, et al. Introns are key regulatory elements of rice tubulin expression[J]. Planta, 2004, 218(5): 693−703. doi: 10.1007/s00425-003-1150-0 [27] 唐立群, 肖层林, 王伟平. SNP分子标记的研究及其应用进展[J]. 中国农学通报, 2012, 28(12): 154−158. doi: 10.11924/j.issn.1000-6850.2012-0074Tang Liqun, Xiao Cenglin, Wang Weiping. Research and application progress of SNP markers[J]. Chinese Agricultural Science Bulletin, 2012, 28(12): 154−158. doi: 10.11924/j.issn.1000-6850.2012-0074 [28] 赵家熙, 崔宝月, 董迎辉, 等. 缢蛏生长因子受体结合蛋白2基因克隆、时空表达及SNP筛查[J]. 海洋学报, 2018, 40(2): 87−94.Zhao Jiaxi, Cui Baoyue, Dong Yinghui, et al. Cloning, spatiotemporal expression and SNPs identification of GRB2 gene in Sinonovacula constricta[J]. Haiyang Xuebao, 2018, 40(2): 87−94. [29] 刘思玮, 李琪, 于红, 等. 长牡蛎糖原磷酸化酶基因SNPs与生长性状和糖原含量的相关性分析[J]. 中国水产科学, 2013, 20(3): 481−489. doi: 10.3724/SP.J.1118.2013.00481Liu Siwei, Li Qi, Yu Hong, et al. Single nucleotide polymorphisms in glycogen phosphorylase gene and their association with growth performance and glycogen content in Pacific oyster Crassostrea gigas[J]. Journal of Fishery Sciences of China, 2013, 20(3): 481−489. doi: 10.3724/SP.J.1118.2013.00481 -

计量
- 文章访问数: 602
- HTML全文浏览量: 185
- PDF下载量: 29
- 被引次数: 0
